

Molecular recognition between ketosynthase and acyl carrier protein domains of the 6-deoxyerythronolide B synthase
- PMID: 21127271
- PMCID: PMC3009775
- DOI: 10.1073/pnas.1014081107
Molecular recognition between ketosynthase and acyl carrier protein domains of the 6-deoxyerythronolide B synthase
Abstract
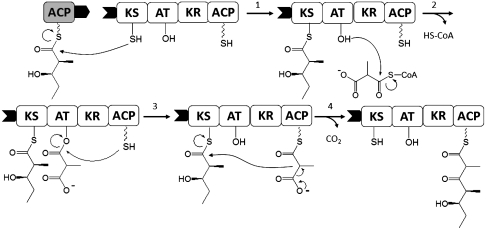
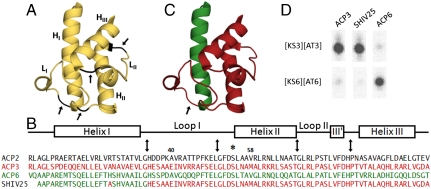
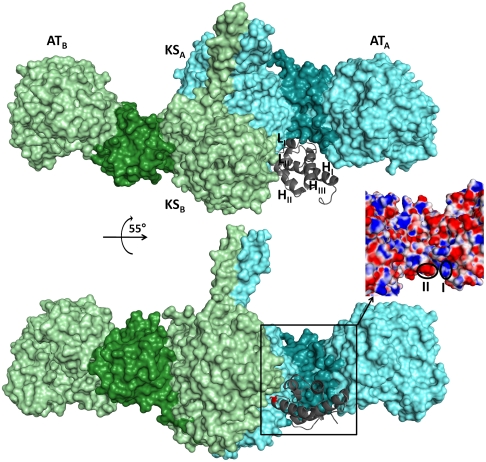
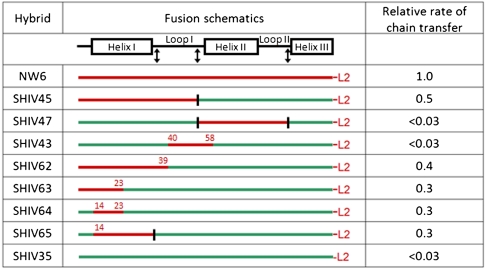
Every polyketide synthase module has an acyl carrier protein (ACP) and a ketosynthase (KS) domain that collaborate to catalyze chain elongation. The same ACP then engages the KS domain of the next module to facilitate chain transfer. Understanding the mechanism for this orderly progress of the growing polyketide chain represents a fundamental challenge in assembly line enzymology. Using both experimental and computational approaches, the molecular basis for KS-ACP interactions in the 6-deoxyerythronolide B synthase has been decoded. Surprisingly, KS-ACP recognition is controlled at different interfaces during chain elongation versus chain transfer. In fact, chain elongation is controlled at a docking site remote from the catalytic center. Not only do our findings reveal a new principle in the modular control of polyketide antibiotic biosynthesis, they also provide a rationale for the mandatory homodimeric structure of polyketide synthases, in contrast to the monomeric nonribosomal peptide synthetases.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
Probing the interactions of an acyl carrier protein domain from the 6-deoxyerythronolide B synthase.Protein Sci. 2011 Jul;20(7):1244-55. doi: 10.1002/pro.652. Protein Sci. 2011. PMID: 21563224 Free PMC article.
-
Extender unit and acyl carrier protein specificity of ketosynthase domains of the 6-deoxyerythronolide B synthase.J Am Chem Soc. 2006 Mar 8;128(9):3067-74. doi: 10.1021/ja058093d. J Am Chem Soc. 2006. PMID: 16506788 Free PMC article.
-
Mutagenesis Supports AlphaFold Prediction of How Modular Polyketide Synthase Acyl Carrier Proteins Dock With Downstream Ketosynthases.Proteins. 2024 Dec;92(12):1375-1384. doi: 10.1002/prot.26733. Epub 2024 Jul 30. Proteins. 2024. PMID: 39078105 Free PMC article.
-
Architecture of the polyketide synthase module: surprises from electron cryo-microscopy.Curr Opin Struct Biol. 2015 Apr;31:9-19. doi: 10.1016/j.sbi.2015.02.014. Epub 2015 Mar 16. Curr Opin Struct Biol. 2015. PMID: 25791608 Free PMC article. Review.
-
Probing the structure and function of acyl carrier proteins to unlock the strategic redesign of type II polyketide biosynthetic pathways.J Biol Chem. 2021 Jan-Jun;296:100328. doi: 10.1016/j.jbc.2021.100328. Epub 2021 Jan 23. J Biol Chem. 2021. PMID: 33493513 Free PMC article. Review.
Cited by
-
In Vitro Reconstitution of Metabolic Pathways: Insights into Nature's Chemical Logic.Synlett. 2015;26(8):1008-1025. doi: 10.1055/s-0034-1380264. Synlett. 2015. PMID: 26207083 Free PMC article.
-
Towards Precision Engineering of Canonical Polyketide Synthase Domains: Recent Advances and Future Prospects.Molecules. 2017 Feb 5;22(2):235. doi: 10.3390/molecules22020235. Molecules. 2017. PMID: 28165430 Free PMC article. Review.
-
Engineered biosynthesis of milbemycins in the avermectin high-producing strain Streptomyces avermitilis.Microb Cell Fact. 2017 Jan 17;16(1):9. doi: 10.1186/s12934-017-0626-8. Microb Cell Fact. 2017. PMID: 28095865 Free PMC article.
-
Engineering the acyltransferase substrate specificity of assembly line polyketide synthases.J R Soc Interface. 2013 May 29;10(85):20130297. doi: 10.1098/rsif.2013.0297. Print 2013 Aug 6. J R Soc Interface. 2013. PMID: 23720536 Free PMC article. Review.
-
Structure and mechanistic analyses of the gating mechanism of elongating ketosynthases.ACS Catal. 2021 Jun 18;11(12):6787-6799. doi: 10.1021/acscatal.1c00745. Epub 2021 May 26. ACS Catal. 2021. PMID: 36187225 Free PMC article.
References
-
- Khosla C, Tang Y, Chen AY, Schnarr NA, Cane DE. Structure and mechanism of the 6-deoxyerythronolide B synthase. Annu Rev Biochem. 2007;76:195–221. - PubMed
-
- Weissman KJ, Leadlay PF. Combinatorial biosynthesis of reduced polyketides. Nat Rev Microbiol. 2005;3:925–936. - PubMed
-
- Wenzel SC, Muller R. Formation of novel secondary metabolites by bacterial multimodular assembly lines: Deviations from textbook biosynthetic logic. Curr Opin Chem Biol. 2005;9:447–458. - PubMed
-
- McDaniel R, Welch M, Hutchinson CR. Genetic approaches to polyketide antibiotics. 1. Chem Rev. 2005;105:543–558. - PubMed
-
- Hill AM. The biosynthesis, molecular genetics, and enzymology of the polyketide-derived metabolites. Nat Prod Rep. 2006;23:256–320. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
