

OCT2 and MATE1 provide bidirectional agmatine transport
- PMID: 21128598
- PMCID: PMC4589871
- DOI: 10.1021/mp100180a
OCT2 and MATE1 provide bidirectional agmatine transport
Abstract
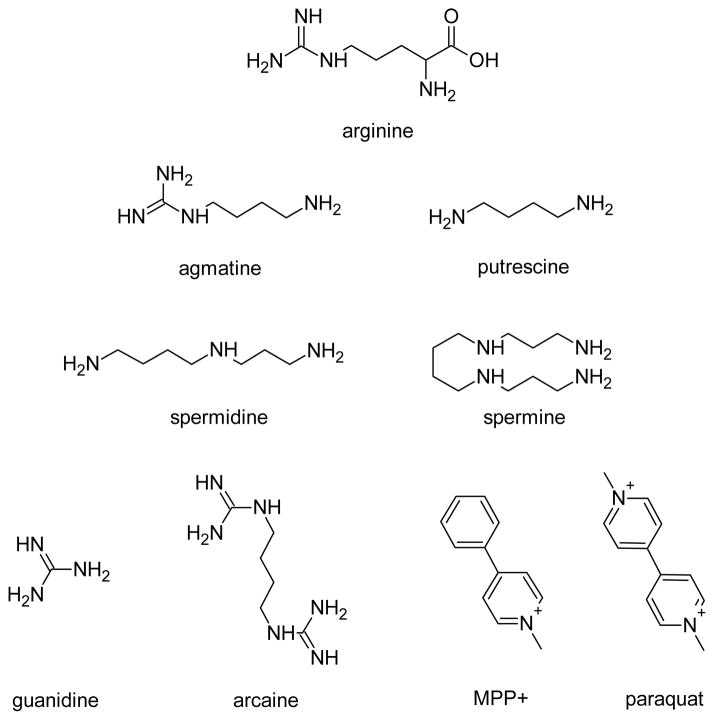
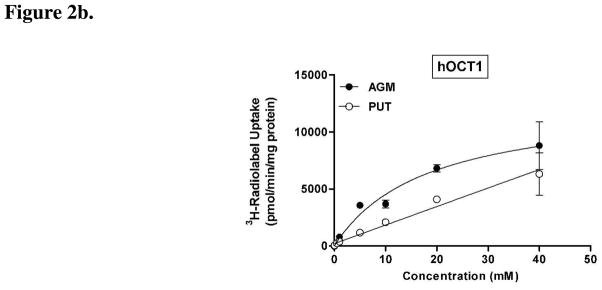
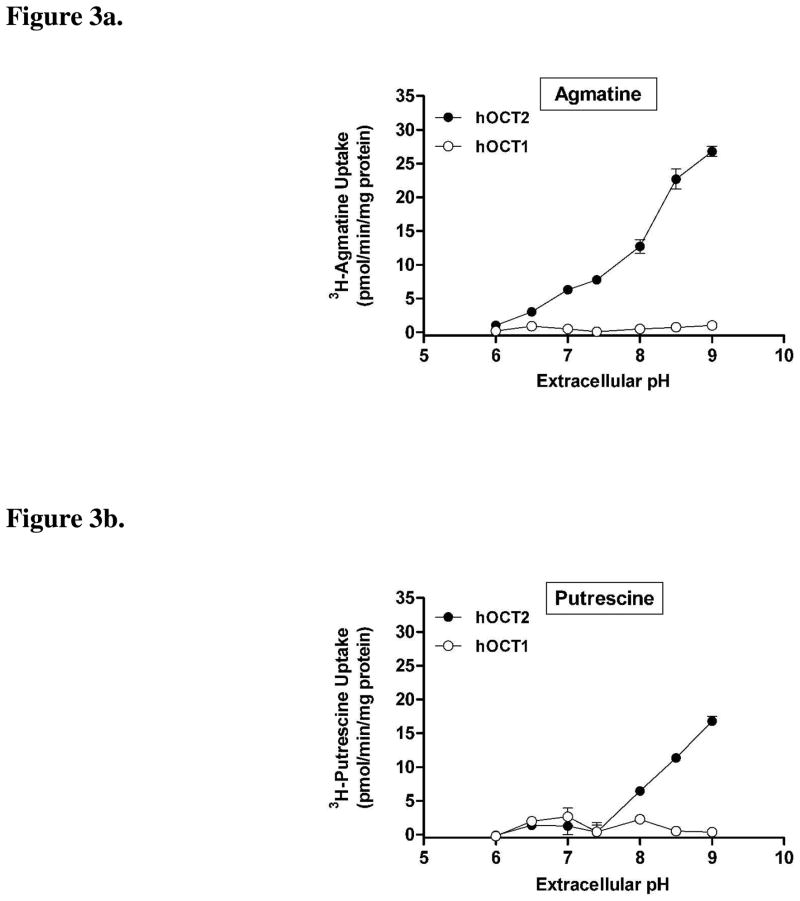
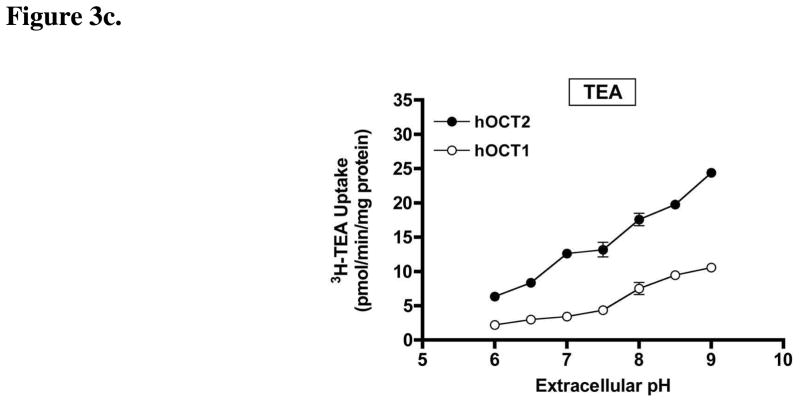
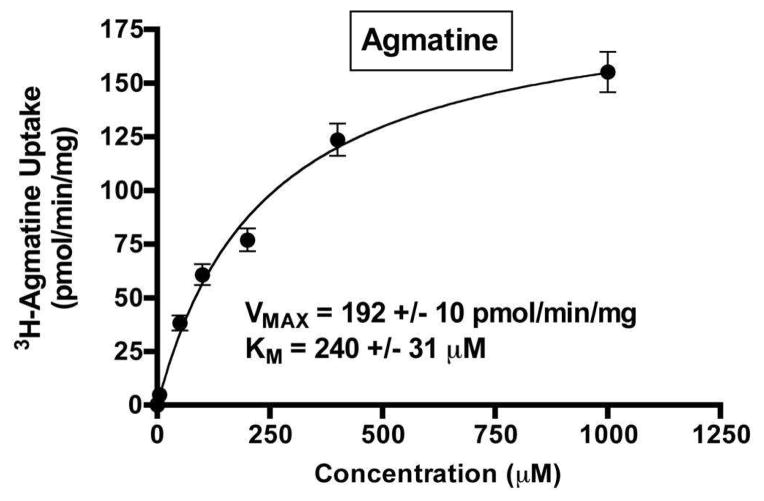
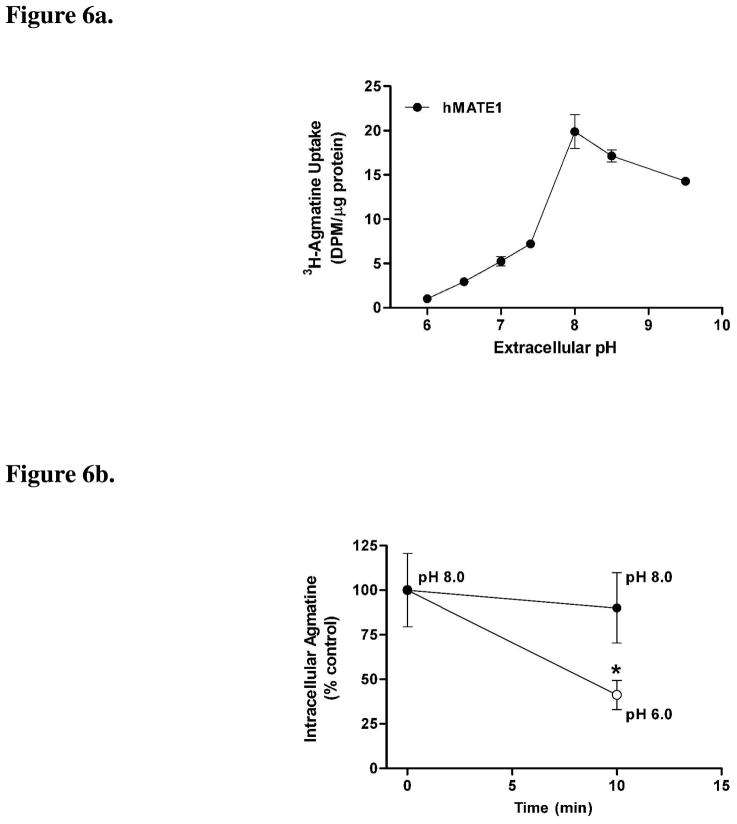
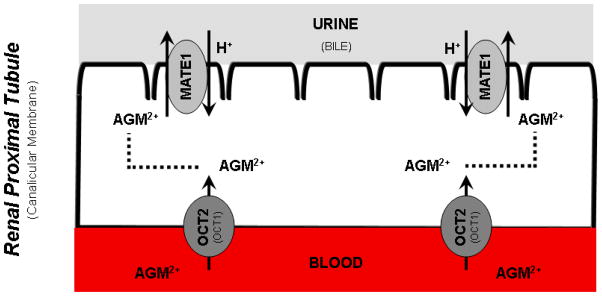
Agmatine is a biogenic amine (l-arginine metabolite) of potential relevance to several central nervous system (CNS) conditions. The identities of transporters underlying agmatine and polyamine disposition in mammalian systems are not well-defined. The SLC-family organic cation transporters (OCT) OCT1 and OCT2 and multidrug and toxin extrusion transporter-1 (MATE1) are transport systems that may be of importance for the cellular disposition of agmatine and putrescine. We investigated the transport of [(3)H]agmatine and [(3)H]putrescine in human embryonic kidney (HEK293) cells stably transfected with hOCT1, hOCT2, and hMATE1. Agmatine transport by hOCT1 and hOCT2 was concentration-dependent, whereas only hOCT2 demonstrated pH-dependent transport. hOCT2 exhibited a greater affinity for agmatine (K(m) = 1.84 ± 0.38 mM) than did hOCT1 (K(m) = 18.73 ± 4.86 mM). Putrescine accumulation was pH- and concentration-dependent in hOCT2-HEK cells (K(m) = 11.29 ± 4.26 mM) but not hOCT1-HEK cells. Agmatine accumulation, in contrast to putrescine, was significantly enhanced by hMATE1 overexpression, and was saturable (K(m) = 240 ± 31 μM; V(max) = 192 ± 10 pmol/min/mg of protein). Intracellular agmatine was also trans-stimulated (effluxed) from hMATE1-HEK cells in the presence of an inward proton-gradient. The hMATE1-mediated transport of agmatine was inhibited by polyamines, the prototypical substrates MPP+ and paraquat, as well as guanidine and arcaine, but not l-arginine. These results suggest that agmatine disposition may be influenced by hOCT2 and hMATE1, two transporters critical in the renal elimination of xenobiotic compounds.
Figures



References
-
- Katsuta H, Takaoka T, Nose K, Nagai Y. Effects of polyamines on the proliferation of mammalian cells in tissue culture. Jpn J Exp Med. 1975;45:345–354. - PubMed
-
- Wolf C, Bruss M, Hanisch B, Gothert M, von Kugelgen I, Molderings GJ. Molecular basis for the antiproliferative effect of agmatine in tumor cells of colonic, hepatic, and neuronal origin. Mol Pharmacol. 2007;71:276–283. - PubMed
-
- Dudkowska M, Lai J, Gardini G, Stachurska A, Grzelakowska-Sztabert B, Colombatto S, Manteuffel-Cymborowska M. Agmatine modulates the in vivo biosynthesis and interconversion of polyamines and cell proliferation. Biochim Biophys Acta. 2003;1619:159–166. - PubMed
-
- Mayeur C, Veuillet G, Michaud M, Raul F, Blottiere HM, Blachier F. Effects of agmatine accumulation in human colon carcinoma cells on polyamine metabolism, DNA synthesis and the cell cycle. Biochim Biophys Acta. 2005;1745:111–123. - PubMed
-
- Satriano J, Matsufuji S, Murakami Y, Lortie MJ, Schwartz D, Kelly CJ, Hayashi S, Blantz RC. Agmatine suppresses proliferation by frameshift induction of antizyme and attenuation of cellular polyamine levels. J Biol Chem. 1998;273:15313–15316. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
