

Parallel reinforcement pathways for conditioned food aversions in the honeybee
- PMID: 21129969
- PMCID: PMC3011020
- DOI: 10.1016/j.cub.2010.11.040
Parallel reinforcement pathways for conditioned food aversions in the honeybee
Abstract
Avoiding toxins in food is as important as obtaining nutrition. Conditioned food aversions have been studied in animals as diverse as nematodes and humans [1, 2], but the neural signaling mechanisms underlying this form of learning have been difficult to pinpoint. Honeybees quickly learn to associate floral cues with food [3], a trait that makes them an excellent model organism for studying the neural mechanisms of learning and memory. Here we show that honeybees not only detect toxins but can also learn to associate odors with both the taste of toxins and the postingestive consequences of consuming them. We found that two distinct monoaminergic pathways mediate learned food aversions in the honeybee. As for other insect species conditioned with salt or electric shock reinforcers [4-7], learned avoidances of odors paired with bad-tasting toxins are mediated by dopamine. Our experiments are the first to identify a second, postingestive pathway for learned olfactory aversions that involves serotonin. This second pathway may represent an ancient mechanism for food aversion learning conserved across animal lineages.
Figures
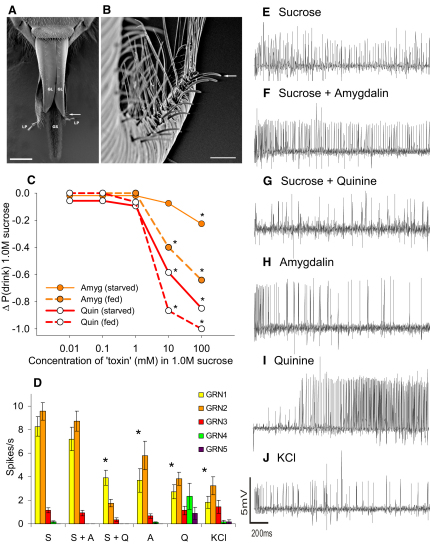
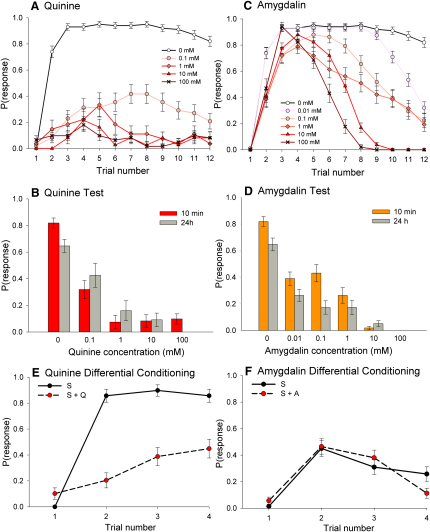
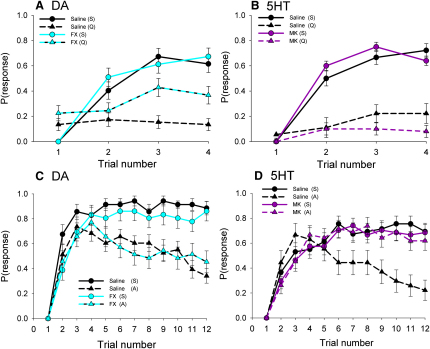
Similar articles
-
The role of dopamine and serotonin in conditioned food aversion learning in the honeybee.Commun Integr Biol. 2011 May;4(3):318-20. doi: 10.4161/cib.4.3.14840. Commun Integr Biol. 2011. PMID: 21980568 Free PMC article.
-
Interactions between conditioned and unconditioned flavor preferences.J Exp Psychol Anim Behav Process. 2005 Oct;31(4):407-17. doi: 10.1037/0097-7403.31.4.407. J Exp Psychol Anim Behav Process. 2005. PMID: 16248727
-
Selective and nonselective serotonin antagonists block the aversive stimulus properties of MK212 and m-chlorophenylpiperazine (mCPP) in mice.Neuropharmacology. 2005 Dec;49(8):1210-9. doi: 10.1016/j.neuropharm.2005.07.015. Epub 2005 Sep 13. Neuropharmacology. 2005. PMID: 16165167
-
Learned flavor preferences. The variable potency of post-oral nutrient reinforcers.Appetite. 2008 Nov;51(3):743-6. doi: 10.1016/j.appet.2008.05.059. Epub 2008 Jul 7. Appetite. 2008. PMID: 18602723 Free PMC article. Review.
-
Acquired aversions as the basis for varied diets of ruminants foraging on rangelands.J Anim Sci. 1996 Aug;74(8):2010-20. doi: 10.2527/1996.7482010x. J Anim Sci. 1996. PMID: 8856457 Review.
Cited by
-
Selenium toxicity to honey bee (Apis mellifera L.) pollinators: effects on behaviors and survival.PLoS One. 2012;7(4):e34137. doi: 10.1371/journal.pone.0034137. Epub 2012 Apr 13. PLoS One. 2012. PMID: 22514621 Free PMC article.
-
Caffeine in floral nectar enhances a pollinator's memory of reward.Science. 2013 Mar 8;339(6124):1202-4. doi: 10.1126/science.1228806. Science. 2013. PMID: 23471406 Free PMC article.
-
A Novel Behavioral Assay to Investigate Gustatory Responses of Individual, Freely-moving Bumble Bees (Bombus terrestris).J Vis Exp. 2016 Jul 21;(113):54233. doi: 10.3791/54233. J Vis Exp. 2016. PMID: 27500630 Free PMC article.
-
Characterization and its implication of a novel taste receptor detecting nutrients in the honey bee, Apis mellifera.Sci Rep. 2019 Aug 12;9(1):11620. doi: 10.1038/s41598-019-46738-z. Sci Rep. 2019. PMID: 31406120 Free PMC article.
-
Differential gene expression analysis following olfactory learning in honeybee (Apis mellifera L.).PLoS One. 2022 Feb 9;17(2):e0262441. doi: 10.1371/journal.pone.0262441. eCollection 2022. PLoS One. 2022. PMID: 35139088 Free PMC article.
References
-
- Reilly S. Central gustatory system lesions and conditioned taste aversion. In: Reilly S., Schachtman T.R., editors. Conditioned Taste Aversion: Behavioral and Neural Processes. Oxford University Press; Oxford: 2009. pp. 309–327.
-
- Zhang Y., Lu H., Bargmann C.I. Pathogenic bacteria induce aversive olfactory learning in Caenorhabditis elegans. Nature. 2005;438:179–184. - PubMed
-
- Bitterman M.E., Menzel R., Fietz A., Schäfer S. Classical conditioning of proboscis extension in honeybees (Apis mellifera) J. Comp. Psychol. 1983;97:107–119. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
