

Production of active recombinant eIF5A: reconstitution in E.coli of eukaryotic hypusine modification of eIF5A by its coexpression with modifying enzymes
- PMID: 21131325
- PMCID: PMC3038461
- DOI: 10.1093/protein/gzq110
Production of active recombinant eIF5A: reconstitution in E.coli of eukaryotic hypusine modification of eIF5A by its coexpression with modifying enzymes
Abstract
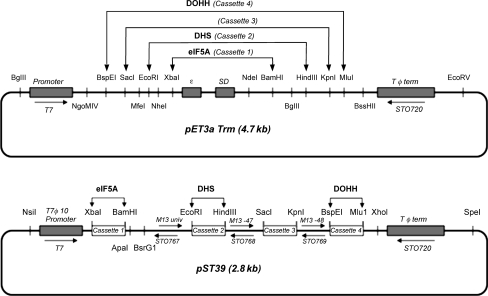
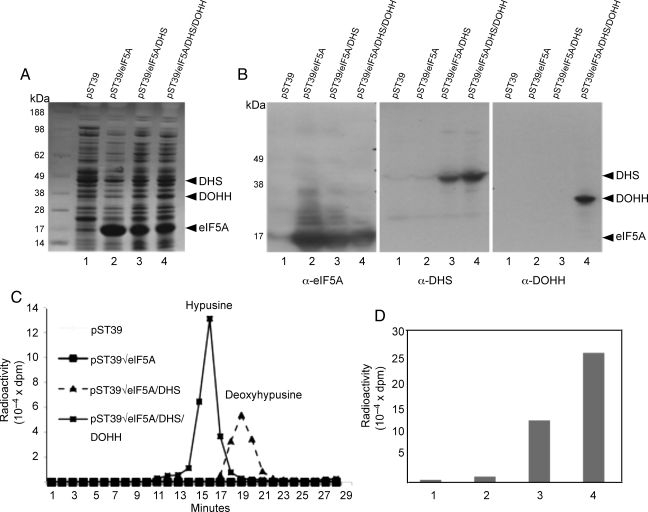
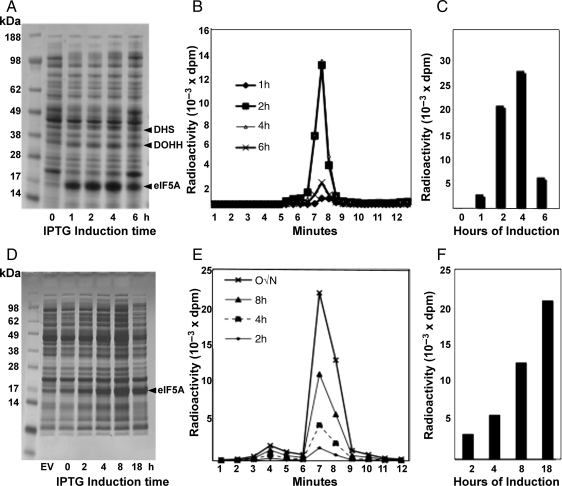
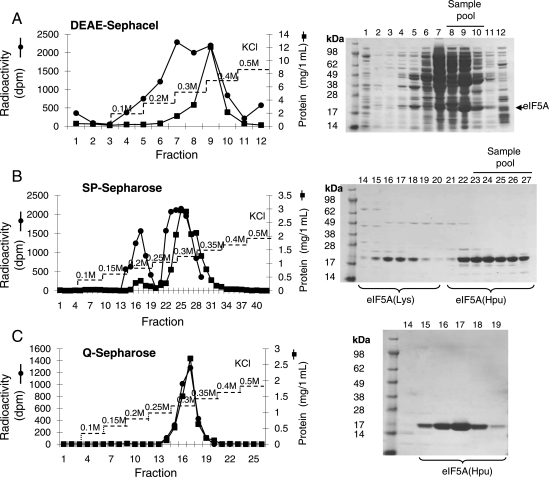
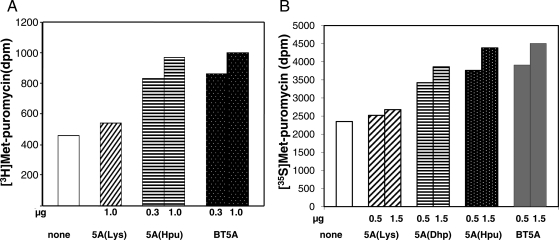
Eukaryotic translation initiation factor 5A (eIF5A) is the only cellular protein that contains the polyamine-modified lysine, hypusine [N(ε)-(4-amino-2-hydroxybutyl)lysine]. Hypusine occurs only in eukaryotes and certain archaea, but not in eubacteria. It is formed post-translationally by two consecutive enzymatic reactions catalyzed by deoxyhypusine synthase (DHS) and deoxyhypusine hydroxylase (DOHH). Hypusine modification is essential for the activity of eIF5A and for eukaryotic cell proliferation. eIF5A binds to the ribosome and stimulates translation in a hypusine-dependent manner, but its mode of action in translation is not well understood. Since quantities of highly pure hypusine-modified eIF5A is desired for structural studies as well as for determination of its binding sites on the ribosome, we have used a polycistronic vector, pST39, to express eIF5A alone, or to co-express human eIF5A-1 with DHS or with both DHS and DOHH in Escherichia coli cells, to engineer recombinant proteins, unmodified eIF5A, deoxyhypusine- or hypusine-modified eIF5A. We have accomplished production of three different forms of recombinant eIF5A in high quantity and purity. The recombinant hypusine-modified eIF5A was as active in methionyl-puromycin synthesis as the native, eIF5A (hypusine form) purified from mammalian tissue. The recombinant eIF5A proteins will be useful tools in future structure/function and the mechanism studies in translation.
Figures
References
-
- Barrios A., Selleck W., Hnatkovich B., Kramer R., Sermwittayawong D., Tan S. Methods. 2007;41:271–277. doi:10.1016/j.ymeth.2006.08.007. - DOI - PMC - PubMed
-
- Benne R., Hershey J.W. J. Biol. Chem. 1978;253:3078–3087. - PubMed
-
- Blaha G., Stanley R.E., Steitz T.A. Science. 2009;325:966–970. doi:10.1126/science.1175800. - DOI - PMC - PubMed
-
- Cano V.S., Jeon G.A., Johansson H.E., Henderson C.A., Park J.H., Valentini S.R., Hershey J.W., Park M.H. FEBS J. 2008;275:44–58. doi:10.1111/j.1742-4658.2007.06172.x. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
