

Transcriptional analysis of a Photorhabdus sp. variant reveals transcriptional control of phenotypic variation and multifactorial pathogenicity in insects
- PMID: 21131515
- PMCID: PMC3028736
- DOI: 10.1128/AEM.01696-10
Transcriptional analysis of a Photorhabdus sp. variant reveals transcriptional control of phenotypic variation and multifactorial pathogenicity in insects
Abstract
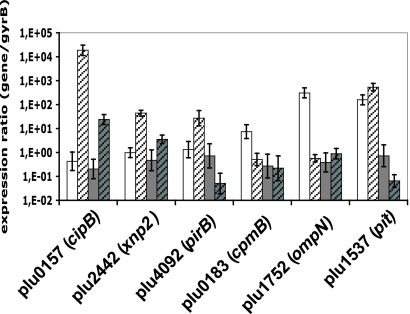
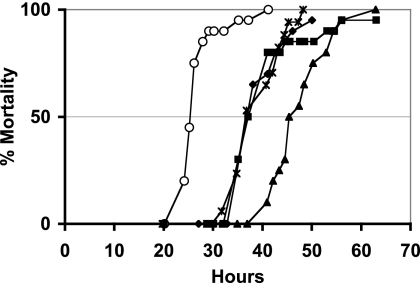
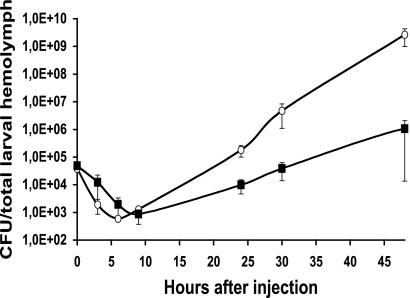
Photorhabdus luminescens lives in a mutualistic association with entomopathogenic nematodes and is pathogenic for insects. Variants of Photorhabdus frequently arise irreversibly and are studied because they have altered phenotypic traits that are potentially important for the host interaction. VAR* is a colonial and phenotypic variant displaying delayed pathogenicity when directly injected into the insect, Spodoptera littoralis. In this study, we evaluated the role of transcriptomic modulation in determining the phenotypic variation and delayed pathogenicity of VAR* with respect to the corresponding wild-type form, TT01α. A P. luminescens microarray identified 148 genes as differentially transcribed between VAR* and TT01α. The net regulator status of VAR* was found to be significantly modified. We also observed in VAR* a decrease in the transcription of genes supporting certain phenotypic traits, such as pigmentation, crystalline inclusion, antibiosis, and protease and lipase activities. Three genes encoding insecticidal toxins (pit and pirB) or putative insecticidal toxins (xnp2) were less transcribed in VAR* than in the TT01α. The overexpression of these genes was not sufficient to restore the virulence of VAR* to the levels of ΤΤ01α, which suggests that the lower virulence of VAR* does not result from impaired toxemia in insects. Three loci involved in oxidative stress responses (sodA, katE, and the hca operon) were found to be downregulated in VAR*. This is consistent with the greater sensitivity of VAR* to H(2)O(2) and may account for the impaired bacteremia in the hemolymph of S. littoralis larvae observed with VAR*. In conclusion, we demonstrate here that some phenotypic traits of VAR* are regulated transcriptionally and highlight the multifactorial nature of pathogenicity in insects.
Figures
Similar articles
-
The HcaR regulatory protein of Photorhabdus luminescens affects the production of proteins involved in oxidative stress and toxemia.Proteomics. 2007 Dec;7(24):4499-510. doi: 10.1002/pmic.200700609. Proteomics. 2007. PMID: 18072208
-
Comparative analysis of the Photorhabdus luminescens and the Yersinia enterocolitica genomes: uncovering candidate genes involved in insect pathogenicity.BMC Genomics. 2008 Jan 25;9:40. doi: 10.1186/1471-2164-9-40. BMC Genomics. 2008. PMID: 18221513 Free PMC article. Review.
-
AstR-AstS, a new two-component signal transduction system, mediates swarming, adaptation to stationary phase and phenotypic variation in Photorhabdus luminescens.Microbiology (Reading). 2004 Apr;150(Pt 4):897-910. doi: 10.1099/mic.0.26563-0. Microbiology (Reading). 2004. PMID: 15073299
-
The PhoP-PhoQ two-component regulatory system of Photorhabdus luminescens is essential for virulence in insects.J Bacteriol. 2004 Mar;186(5):1270-9. doi: 10.1128/JB.186.5.1270-1279.2004. J Bacteriol. 2004. PMID: 14973084 Free PMC article.
-
Regulation of Phenotypic Switching and Heterogeneity in Photorhabdus luminescens Cell Populations.J Mol Biol. 2019 Nov 22;431(23):4559-4568. doi: 10.1016/j.jmb.2019.04.015. Epub 2019 Apr 22. J Mol Biol. 2019. PMID: 31022406 Review.
Cited by
-
Diverse Roles for a Conserved DNA-Methyltransferase in the Entomopathogenic Bacterium Xenorhabdus.Int J Mol Sci. 2022 Oct 9;23(19):11981. doi: 10.3390/ijms231911981. Int J Mol Sci. 2022. PMID: 36233296 Free PMC article.
-
DNA Adenine Methyltransferase (Dam) Overexpression Impairs Photorhabdus luminescens Motility and Virulence.Front Microbiol. 2017 Sep 1;8:1671. doi: 10.3389/fmicb.2017.01671. eCollection 2017. Front Microbiol. 2017. PMID: 28919886 Free PMC article.
-
Nematode-bacterium symbioses--cooperation and conflict revealed in the "omics" age.Biol Bull. 2012 Aug;223(1):85-102. doi: 10.1086/BBLv223n1p85. Biol Bull. 2012. PMID: 22983035 Free PMC article. Review.
-
An antimicrobial peptide-resistant minor subpopulation of Photorhabdus luminescens is responsible for virulence.Sci Rep. 2017 Mar 2;7:43670. doi: 10.1038/srep43670. Sci Rep. 2017. PMID: 28252016 Free PMC article.
-
Protease S of entomopathogenic bacterium Photorhabdus laumondii: expression, purification and effect on greater wax moth Galleria mellonella.Mol Biol Rep. 2024 Jun 1;51(1):713. doi: 10.1007/s11033-024-09654-8. Mol Biol Rep. 2024. PMID: 38824247
References
-
- Akhurst, R. J. 1980. Morphological and functional dimorphism in Xenorhabdus spp., bacteria symbiotically associated with the insect pathogenic nematodes Neoaplectana and Heterorhabditis. J. Gen. Microbiol. 121:303-309. - PubMed
-
- Ausubel, F. M., et al. 1993. Current protocols in molecular biology. John Wiley & Sons, Inc., New York, NY.
-
- Beier, D., and R. Gross. 2006. Regulation of bacterial virulence by two-component systems. Curr. Opin. Microbiol. 9:143-152. - PubMed
-
- Boemare, N., J.-O. Thaler, and A. Lanois. 1997. Simple bacteriological tests for phenotypic characterization of Xenorhabdus and Photorhabdus phase variants. Symbiosis 22:167-175.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
