

Japanese quail as a model system for studying the neuroendocrine control of reproductive and social behaviors
- PMID: 21131709
- PMCID: PMC3522869
- DOI: 10.1093/ilar.51.4.310
Japanese quail as a model system for studying the neuroendocrine control of reproductive and social behaviors
Abstract
Japanese quail (Coturnix japonica; referred to simply as quail in this article) readily exhibit sexual behavior and related social behaviors in captive conditions and have therefore proven valuable for studies of how early social experience can shape adult mate preference and sexual behavior. Quail have also been used in sexual conditioning studies illustrating that natural stimuli predict successful reproduction via Pavlovian processes. In addition, they have proven to be a good model to study how variation in photoperiod regulates reproduction and how variation in gonadal steroid hormones controls sexual behavior. For example, studies have shown that testosterone activates male-typical behaviors after being metabolized into estrogenic and androgenic metabolites. A critical site of action for these metabolites is the medial preoptic nucleus (POM), which is larger in males than in females. The enzyme aromatase converts testosterone to estradiol and is enriched in the POM in a male-biased fashion. Quail studies were the first to show that this enzyme is regulated both relatively slowly via genomic actions of steroids and more quickly via phosphorylation. With this base of knowledge and the recent cloning of the entire genome of the closely related chicken, quail will be valuable for future studies connecting gene expression to sexual and social behaviors.
Figures
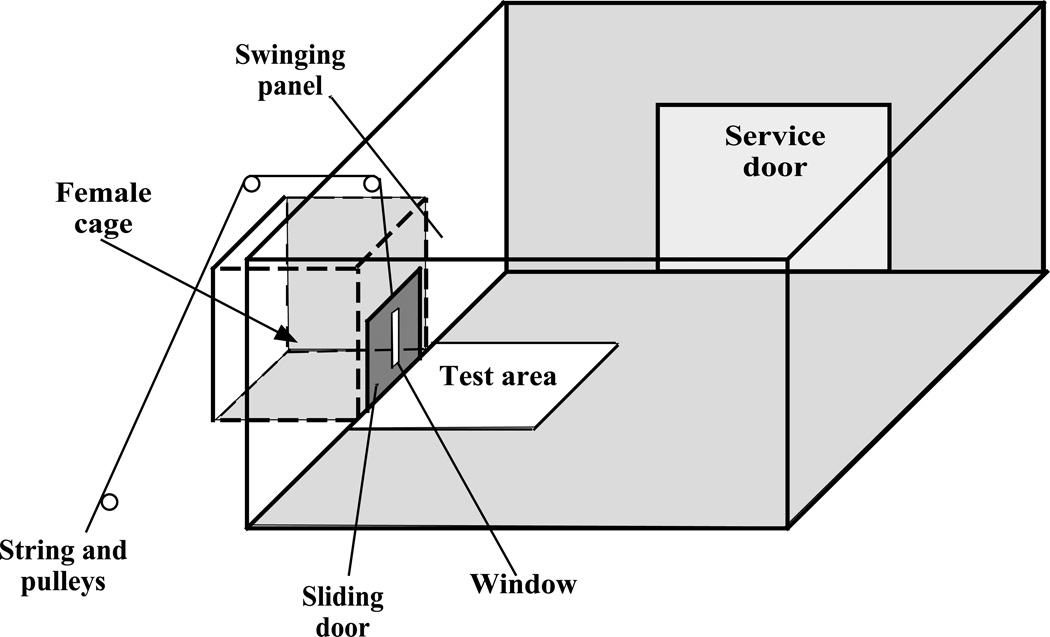
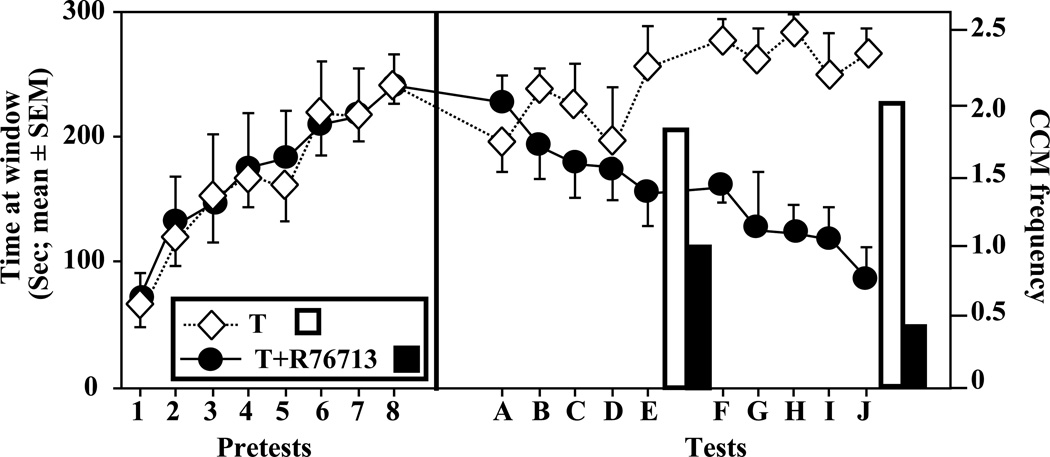
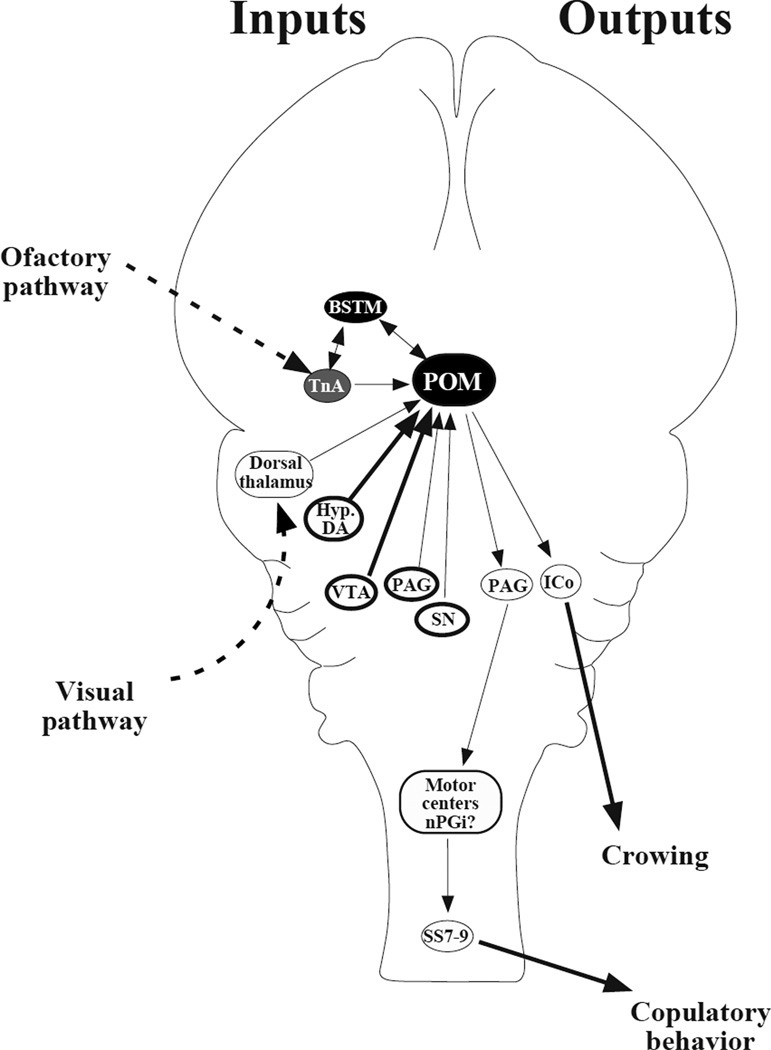
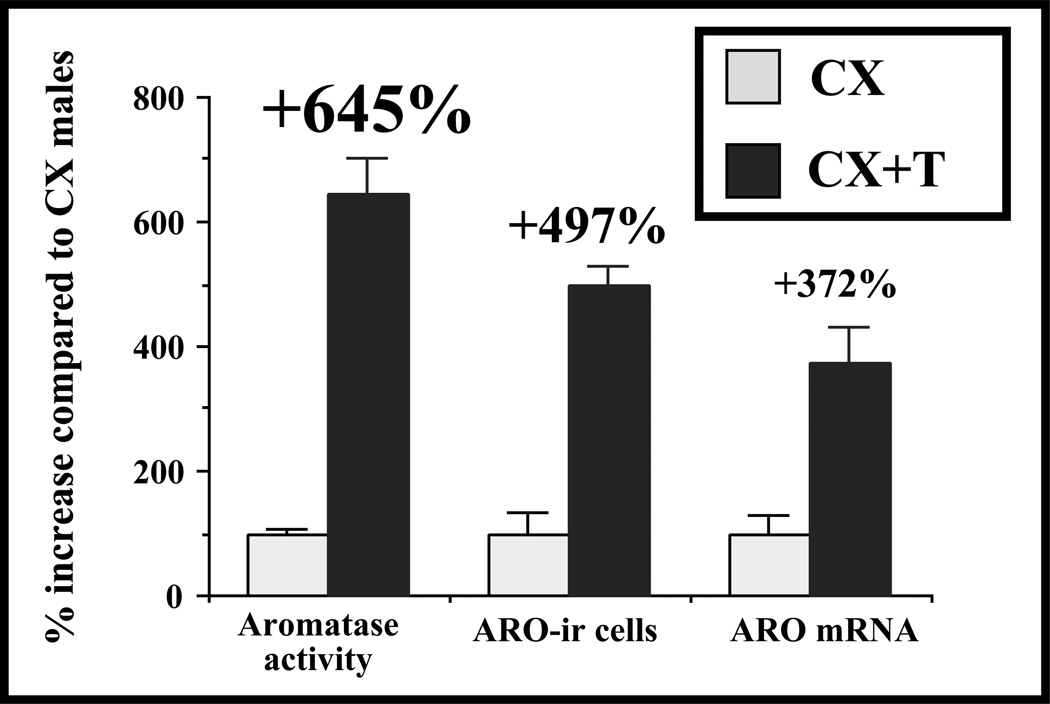

References
-
- Absil P, Baillien M, Ball GF, Panzica G, Balthazart J. The control of preoptic aromatase activity by afferent inputs in Japanese quail. Brain Res Rev. 2001a;37:38–58. - PubMed
-
- Absil P, Riters LV, Balthazart J. Preoptic aromatase cells project to the mesencephalic central gray in the male Japanese quail (Coturnix japonica) Horm Behav. 2001b;40:369–383. - PubMed
-
- Adkins EK. Functional castration of the female Japanese quail. Physiol Behav. 1973;10:619–621. - PubMed
-
- Adkins EK. Hormonal basis of sexual differentiation in the Japanese quail. J Comp Physiol Psychol. 1975;89:61–71. - PubMed
-
- Adkins EK. Effects of diverse androgens on the sexual behavior and morphology of castrated male quail. Horm Behav. 1977;8:201–207. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
