

Nanometer propagation of millisecond motions in V-type allostery
- PMID: 21134639
- PMCID: PMC3003306
- DOI: 10.1016/j.str.2010.09.020
Nanometer propagation of millisecond motions in V-type allostery
Abstract
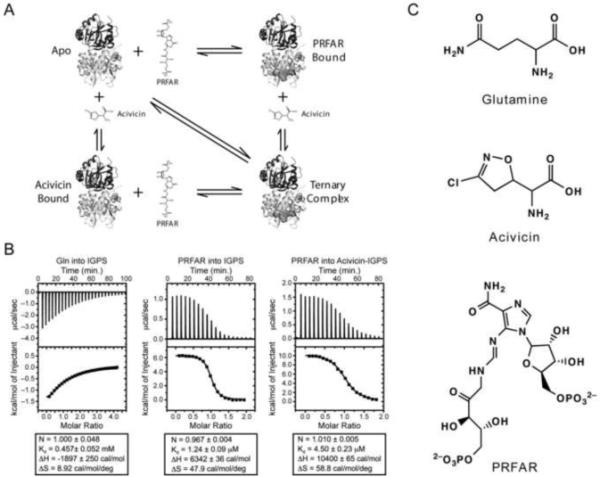
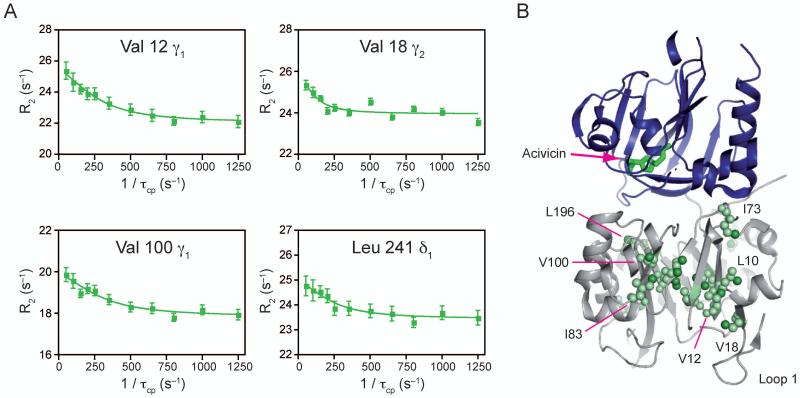
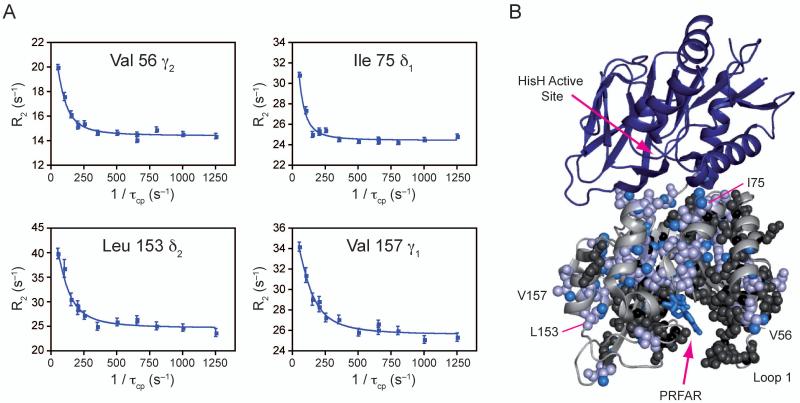
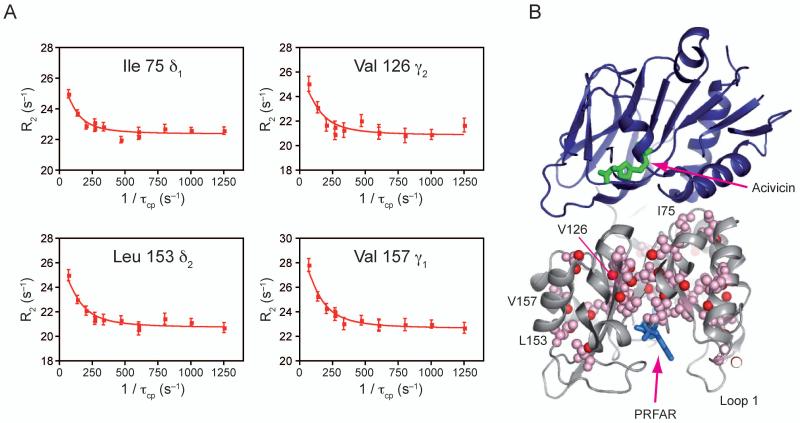
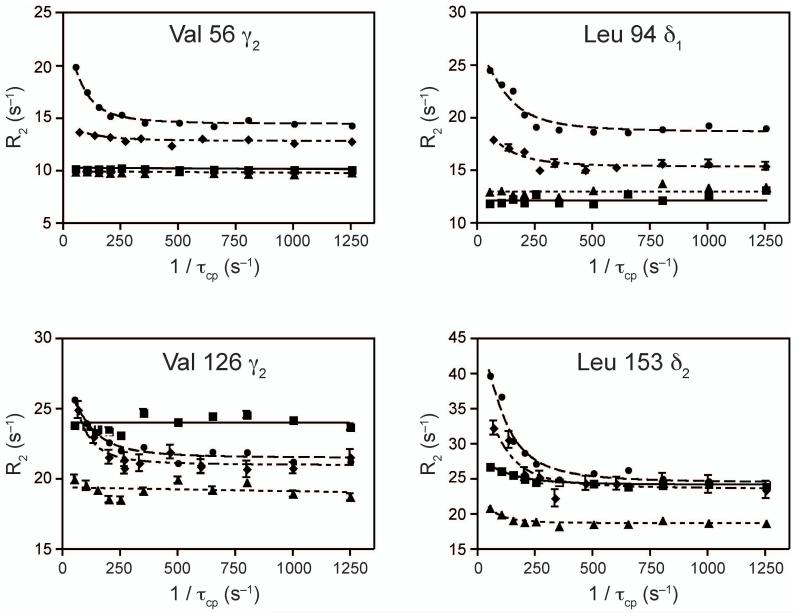
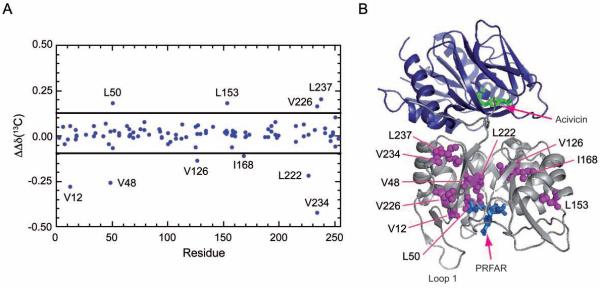
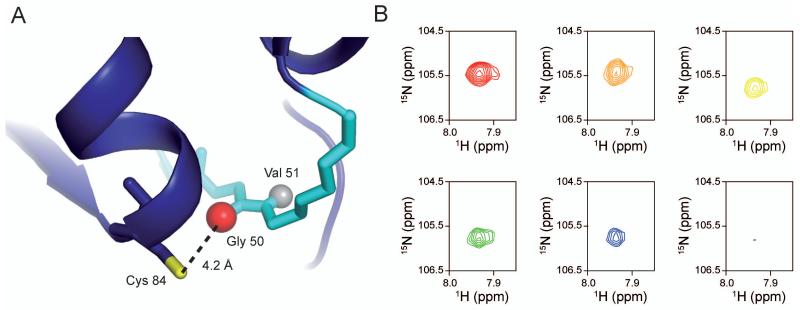
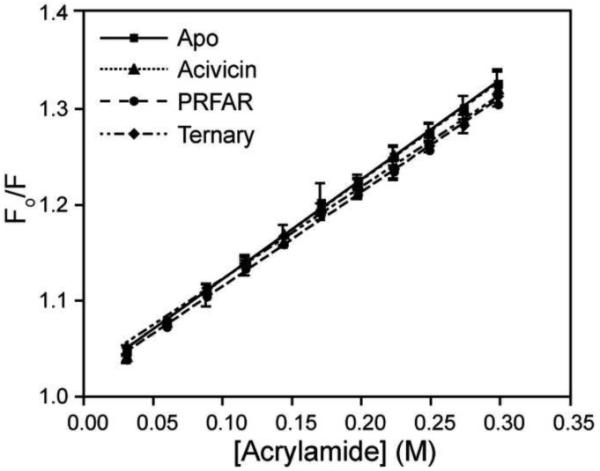
Imidazole glycerol phosphate synthase (IGPS) is a V-type allosteric enzyme, which is catalytically inactive for glutamine hydrolysis until the allosteric effector, N'-[(5'-phosphoribulosyl)formimino]-5-aminoimidazole-4-carboxamide-ribonucleotide (PRFAR) binds 30 Å away. In the apo state, NMR relaxation dispersion experiments indicate the absence of millisecond (ms) timescale motions. Binding of the PRFAR to form the active ternary complex is endothermic with a large positive entropy change. In addition, there is a protein wide enhancement of conformational motions in the ternary complex, which connect the two active sites. NMR chemical shift changes and acrylamide quenching experiments suggest that little in the way of structural changes accompany these motions. The data indicate that enzyme activation in the ternary complex is primarily due to an enhancement of ms motions that allows formation of a population of enzymatically active conformers.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures


References
-
- Ackers GK, Doyle ML, Myers D, Daugherty MA. Molecular code for cooperativity in hemoglobin. Science. 1991;255:54–63. - PubMed
-
- Amaro RE, Sethi A, Myers RS, Davisson VJ, Luthey-Schulten ZA. A network of conserved interactions regulates the allosteric signal in a glutamine amidotransferase. Biochemistry. 2007;46:2156–2173. - PubMed
-
- Beismann-Driemeyer S, Sterner R. Imidazole glycerol phosphate synthase from Thermotoga maritima - Quaternary structure, steady-state kinetics, and reaction mechanism of the bienzyme complex. Journal of Biological Chemistry. 2001;276:20387–20396. - PubMed
-
- Bertrand D, Gopalakrishnan M. Allosteric modulation of nicotinic acetylcholine receptors. Biochem Pharmacol. 2007;74:1155–1163. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
