

Quaternary structure and functional unit of energy coupling factor (ECF)-type transporters
- PMID: 21135102
- PMCID: PMC3037660
- DOI: 10.1074/jbc.M110.199224
Quaternary structure and functional unit of energy coupling factor (ECF)-type transporters
Abstract
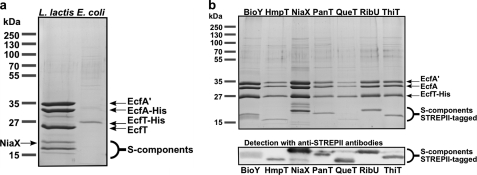
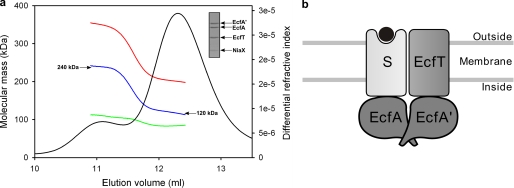
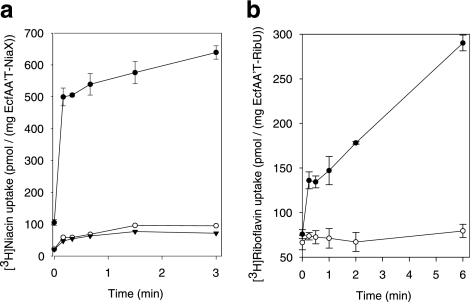
ATP-binding cassette (ABC) transporters mediate transport of diverse substrates across membranes. We have determined the quaternary structure and functional unit of the recently discovered ECF-type (energy coupling factor) of ABC transporters, which is widespread among prokaryotes. ECF transporters are protein complexes consisting of a conserved energizing module (two peripheral ATPases and the integral membrane protein EcfT) and a non-conserved integral membrane protein responsible for substrate specificity (S-component). S-components for different substrates are often unrelated in amino acid sequence but may associate with the same energizing module. Here, the energizing module from Lactococcus lactis was shown to form stable complexes with each of the eight predicted S-components found in the organism. The quaternary structures of three of these complexes were determined by light scattering. EcfT, the two ATPases (EcfA and EcfA'), and the S-components were found to be present in a 1:1:1:1 ratio. The complexes were reconstituted in proteoliposomes and shown to mediate ATP-dependent transport. ECF-type transporters are the smallest known ABC transporters.
Figures
References
-
- Higgins C. F. (1992) Annu. Rev. Cell Biol. 8, 67–113 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
