

Mechanism for the inhibition of the carboxyltransferase domain of acetyl-coenzyme A carboxylase by pinoxaden
- PMID: 21135213
- PMCID: PMC3009786
- DOI: 10.1073/pnas.1012039107
Mechanism for the inhibition of the carboxyltransferase domain of acetyl-coenzyme A carboxylase by pinoxaden
Abstract
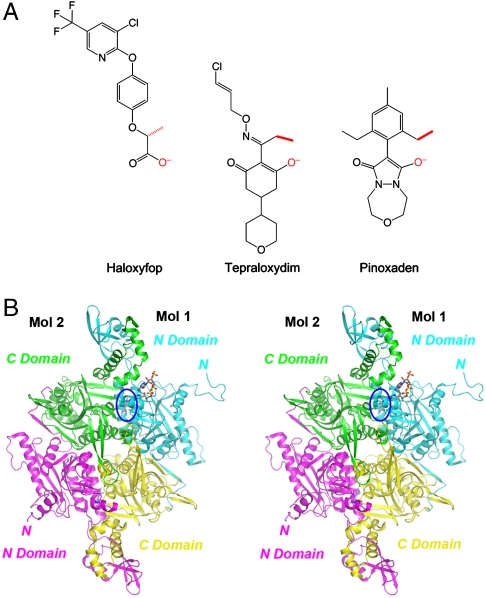
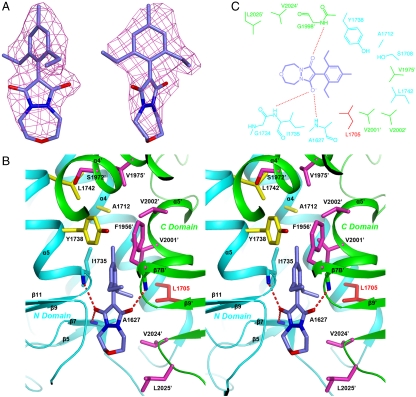
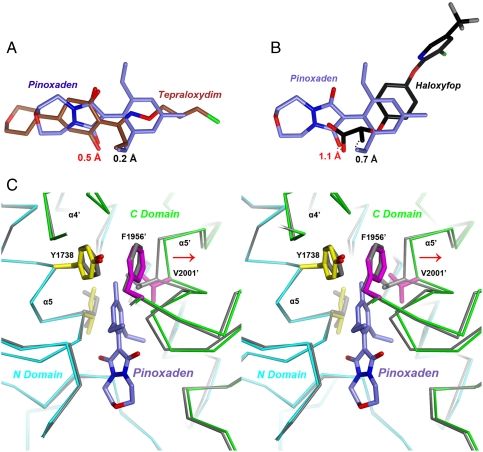
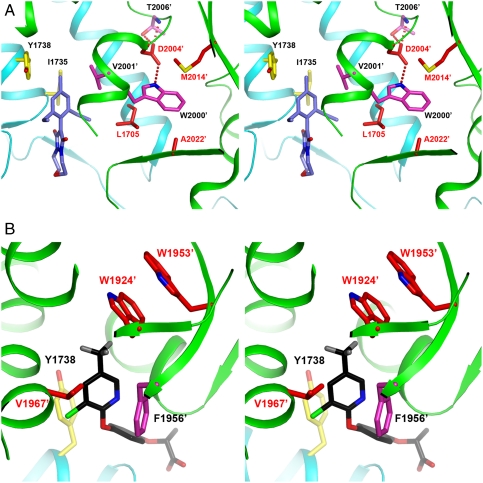
Acetyl-CoA carboxylases (ACCs) are crucial metabolic enzymes and have been targeted for drug development against obesity, diabetes, and other diseases. The carboxyltransferase (CT) domain of this enzyme is the site of action for three different classes of herbicides, as represented by haloxyfop, tepraloxydim, and pinoxaden. Our earlier studies have demonstrated that haloxyfop and tepraloxydim bind in the CT active site at the interface of its dimer. However, the two compounds probe distinct regions of the dimer interface, sharing primarily only two common anchoring points of interaction with the enzyme. We report here the crystal structure of the CT domain of yeast ACC in complex with pinoxaden at 2.8-Å resolution. Despite their chemical diversity, pinoxaden has a similar binding mode as tepraloxydim and requires a small conformational change in the dimer interface for binding. Crystal structures of the CT domain in complex with all three classes of herbicides confirm the importance of the two anchoring points for herbicide binding. The structures also provide a foundation for understanding the molecular basis of the herbicide resistance mutations and cross resistance among the herbicides, as well as for the design and development of new inhibitors against plant and human ACCs.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Wakil SJ, Stoops JK, Joshi VC. Fatty acid synthesis and its regulation. Annu Rev Biochem. 1983;52:537–579. - PubMed
-
- McGarry JD, Brown NF. The mitochondrial carnitine palmitoyltransferase system. From concept to molecular analysis. Eur J Biochem. 1997;244:1–14. - PubMed
-
- Ramsay RR, Gandour RD, van der Leij FR. Molecular enzymology of carnitine transfer and transport. Biochim Biophys Acta. 2001;1546:21–43. - PubMed
-
- Abu-Elheiga L, Matzuk MM, Abo-Hashema KAH, Wakil SJ. Continuous fatty acid oxidation and reduced fat storage in mice lacking acetyl-CoA carboxylase 2. Science. 2001;291:2613–2616. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
