Staphylococcus epidermidis surfactant peptides promote biofilm maturation and dissemination of biofilm-associated infection in mice
- PMID: 21135501
- PMCID: PMC3007140
- DOI: 10.1172/JCI42520
Staphylococcus epidermidis surfactant peptides promote biofilm maturation and dissemination of biofilm-associated infection in mice
Abstract
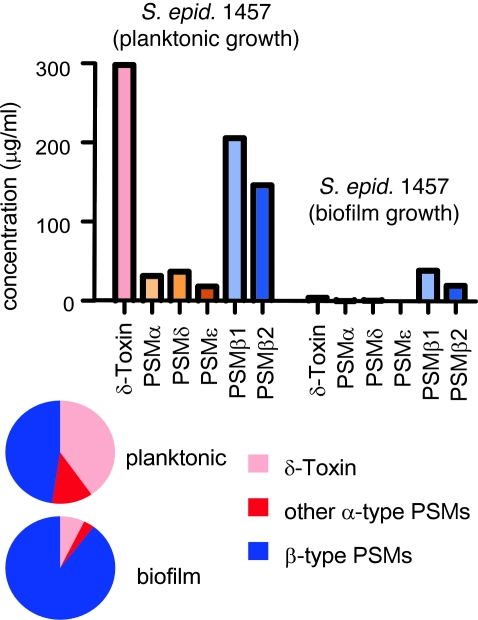
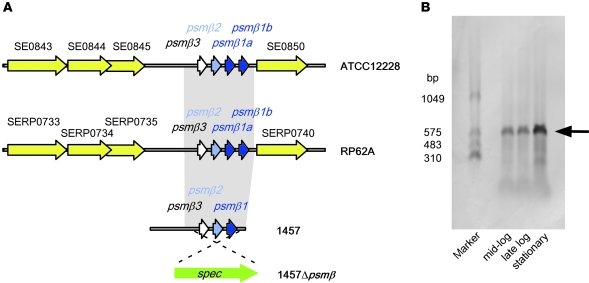
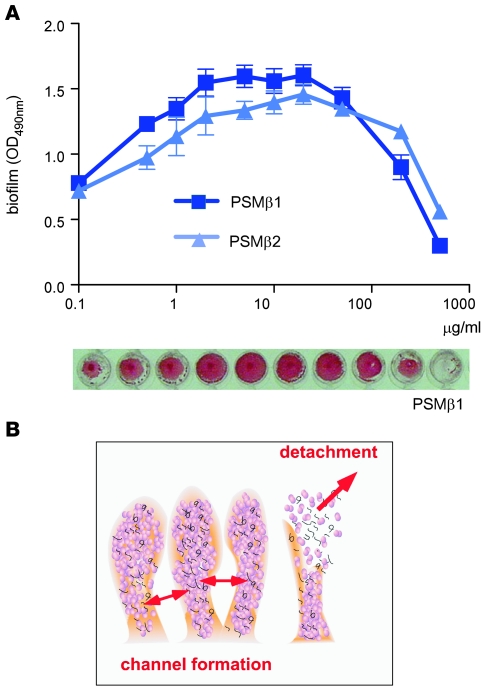
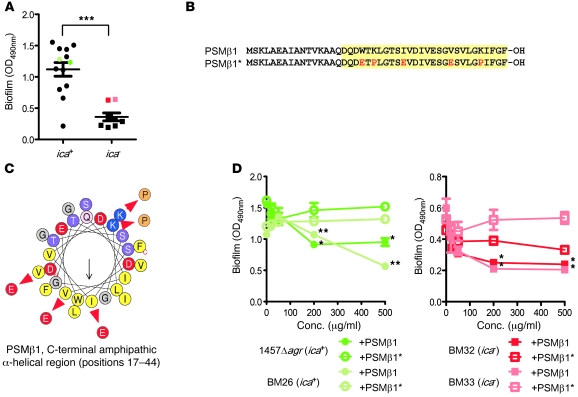
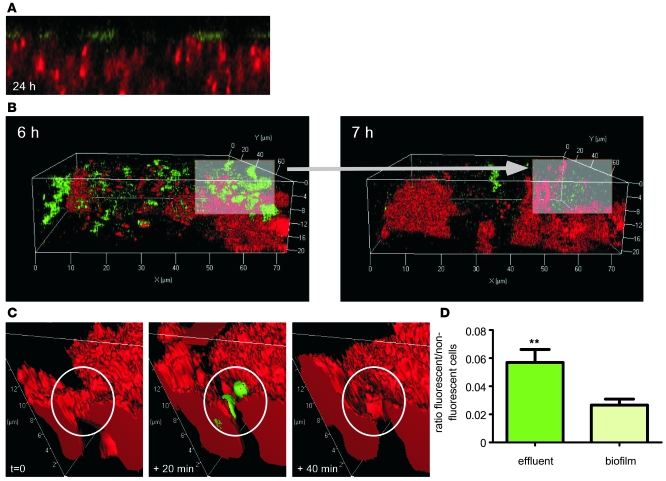
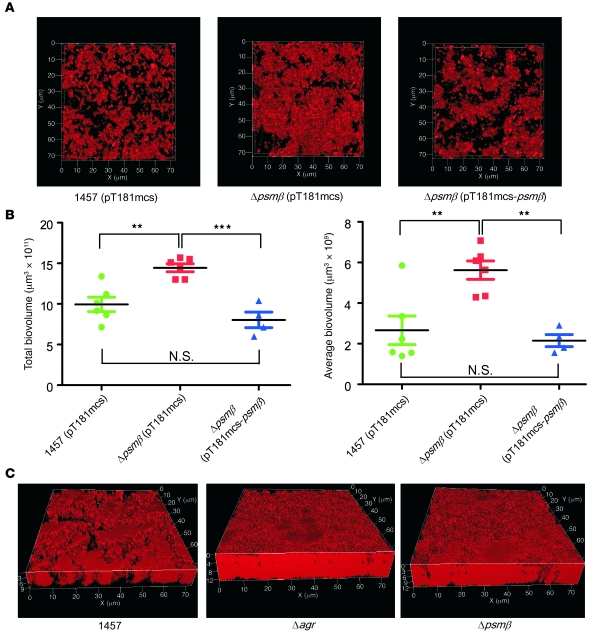
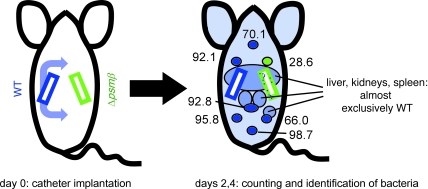
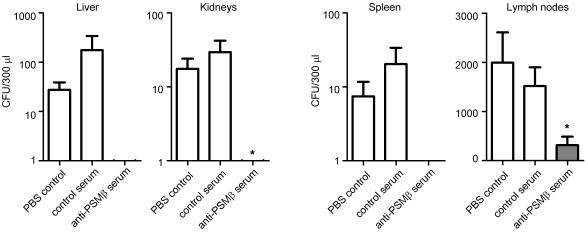
Biofilms are surface-attached agglomerations of microorganisms embedded in an extracellular matrix. Biofilm-associated infections are difficult to eradicate and represent a significant reservoir for disseminating and recurring serious infections. Infections involving biofilms frequently develop on indwelling medical devices in hospitalized patients, and Staphylococcus epidermidis is the leading cause of infection in this setting. However, the molecular determinants of biofilm dissemination are unknown. Here we have demonstrated that specific secreted, surfactant-like S. epidermidis peptides--the β subclass of phenol-soluble modulins (PSMs)--promote S. epidermidis biofilm structuring and detachment in vitro and dissemination from colonized catheters in a mouse model of device-related infection. Our study establishes in vivo significance of biofilm detachment mechanisms for the systemic spread of biofilm-associated infection and identifies the effectors of biofilm maturation and detachment in a premier biofilm-forming pathogen. Furthermore, by demonstrating that antibodies against PSMβ peptides inhibited bacterial spread from indwelling medical devices, we have provided proof of principle that interfering with biofilm detachment mechanisms may prevent dissemination of biofilm-associated infection.
Figures