

Aquaporin3 is a sperm water channel essential for postcopulatory sperm osmoadaptation and migration
- PMID: 21135872
- PMCID: PMC3343308
- DOI: 10.1038/cr.2010.169
Aquaporin3 is a sperm water channel essential for postcopulatory sperm osmoadaptation and migration
Abstract
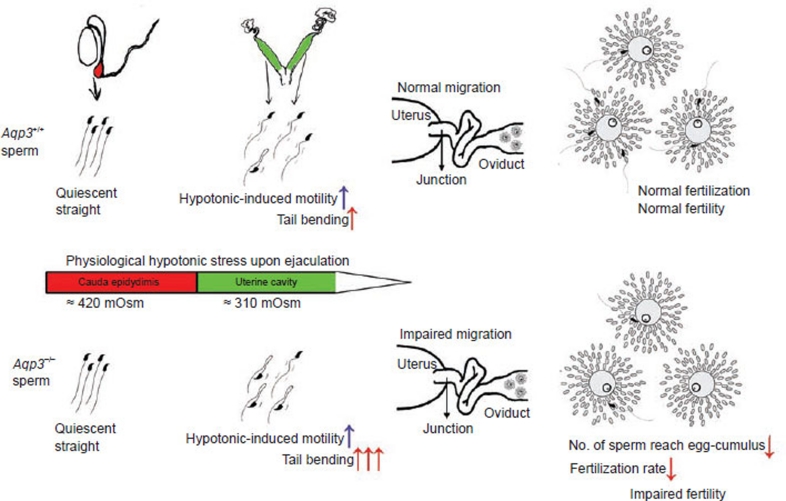
In the journey from the male to female reproductive tract, mammalian sperm experience a natural osmotic decrease (e.g., in mouse, from ~415 mOsm in the cauda epididymis to ~310 mOsm in the uterine cavity). Sperm have evolved to utilize this hypotonic exposure for motility activation, meanwhile efficiently silence the negative impact of hypotonic cell swelling. Previous physiological and pharmacological studies have shown that ion channel-controlled water influx/efflux is actively involved in the process of sperm volume regulation; however, no specific sperm proteins have been found responsible for this rapid osmoadaptation. Here, we report that aquaporin3 (AQP3) is a sperm water channel in mice and humans. Aqp3-deficient sperm show normal motility activation in response to hypotonicity but display increased vulnerability to hypotonic cell swelling, characterized by increased tail bending after entering uterus. The sperm defect is a result of impaired sperm volume regulation and progressive cell swelling in response to physiological hypotonic stress during male-female reproductive tract transition. Time-lapse imaging revealed that the cell volume expansion begins at cytoplasmic droplet, forcing the tail to angulate and form a hairpin-like structure due to mechanical membrane stretch. The tail deformation hampered sperm migration into oviduct, resulting in impaired fertilization and reduced male fertility. These data suggest AQP3 as an essential membrane pathway for sperm regulatory volume decrease (RVD) that balances the "trade-off" between sperm motility and cell swelling upon physiological hypotonicity, thereby optimizing postcopulatory sperm behavior.
Figures
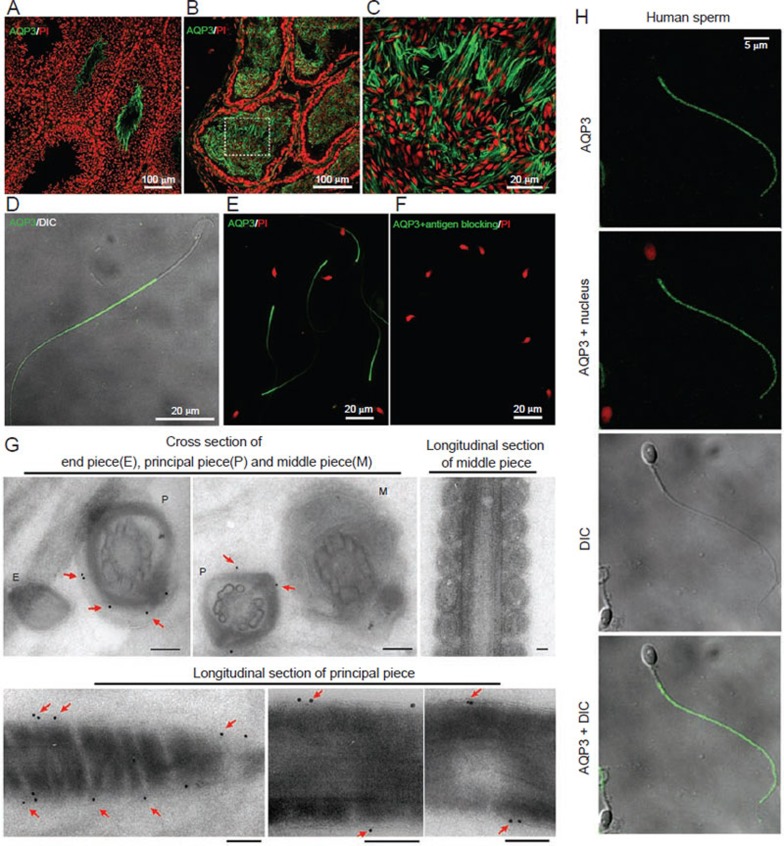
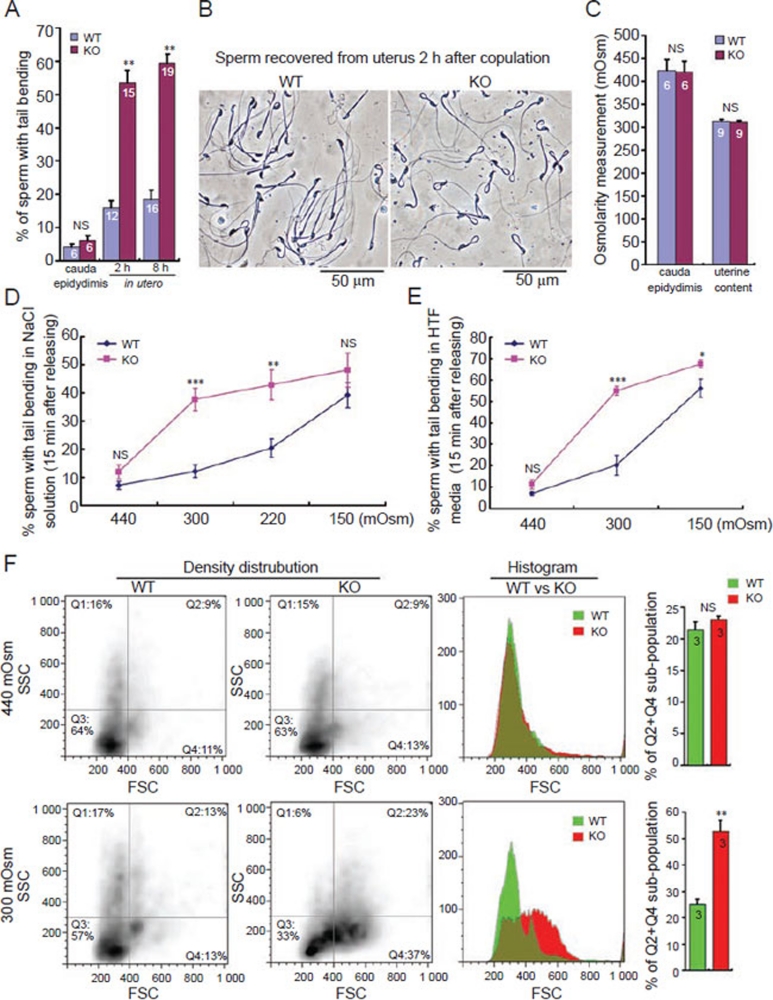
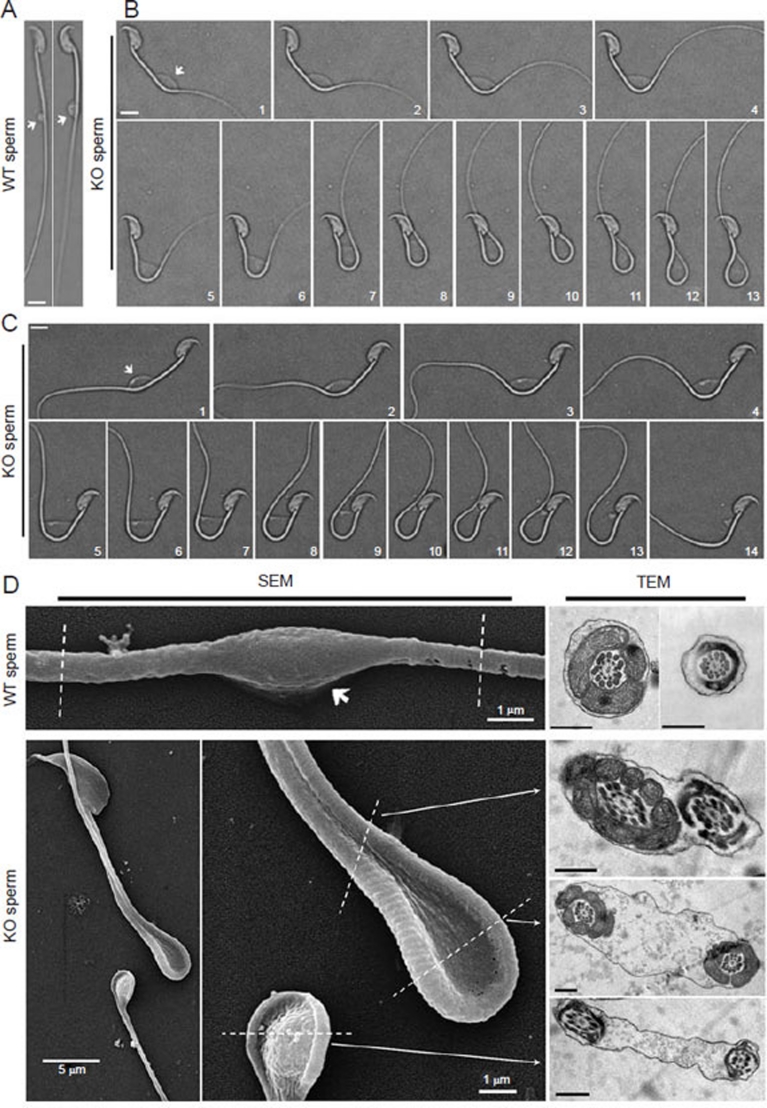
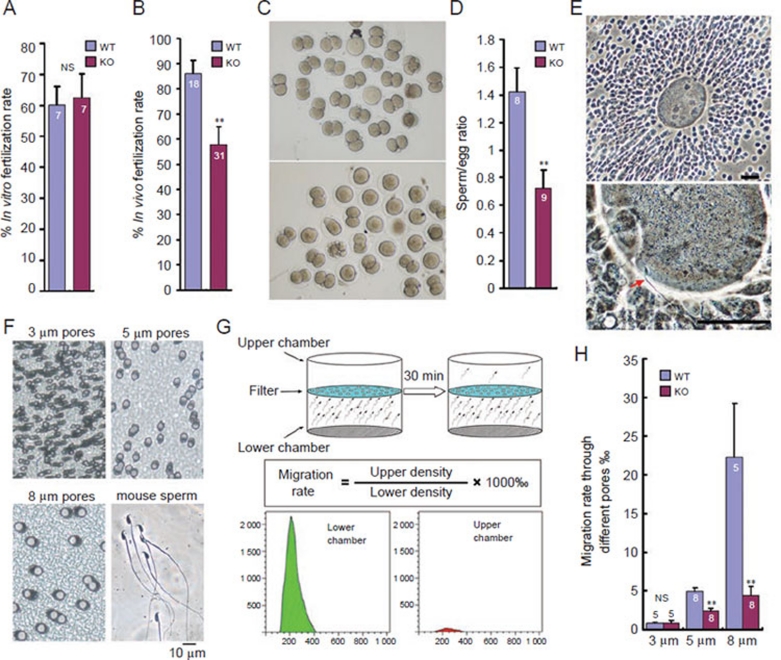
References
-
- Morisawa M, Suzuki K. Osmolality and potassium ion: their roles in initiation of sperm motility in teleosts. Science. 1980;210:1145–1147. - PubMed
-
- Cosson J, Groison AL, Suquet M, et al. Marine fish spermatozoa: racing ephemeral swimmers. Reproduction. 2008;136:277–294. - PubMed
-
- Johnson AL, Howards SS. Hyperosmolality in intraluminal fluids from hamster testis and epididymis: a micropuncture study. Science. 1977;195:492–493. - PubMed
-
- Cooper TG, Yeung CH. Acquisition of volume regulatory response of sperm upon maturation in the epididymis and the role of the cytoplasmic droplet. Microsc Res Tech. 2003;61:28–38. - PubMed
-
- Rossato M, Di Virgilio F, Foresta C. Involvement of osmo-sensitive calcium influx in human sperm activation. Mol Hum Reprod. 1996;2:903–909. - PubMed
