

Regulation and function of TPL-2, an IκB kinase-regulated MAP kinase kinase kinase
- PMID: 21135874
- PMCID: PMC3193413
- DOI: 10.1038/cr.2010.173
Regulation and function of TPL-2, an IκB kinase-regulated MAP kinase kinase kinase
Erratum in
- Cell Res. 2011 Apr;21(4):704
Abstract
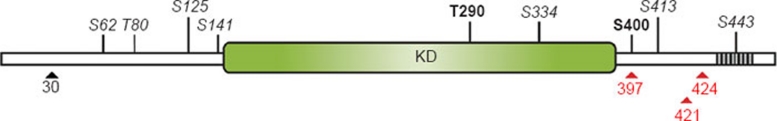
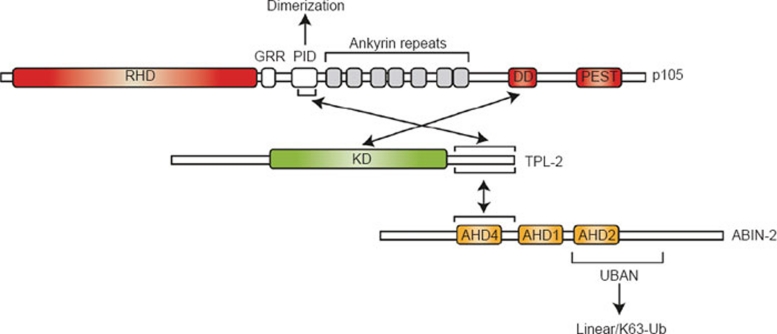
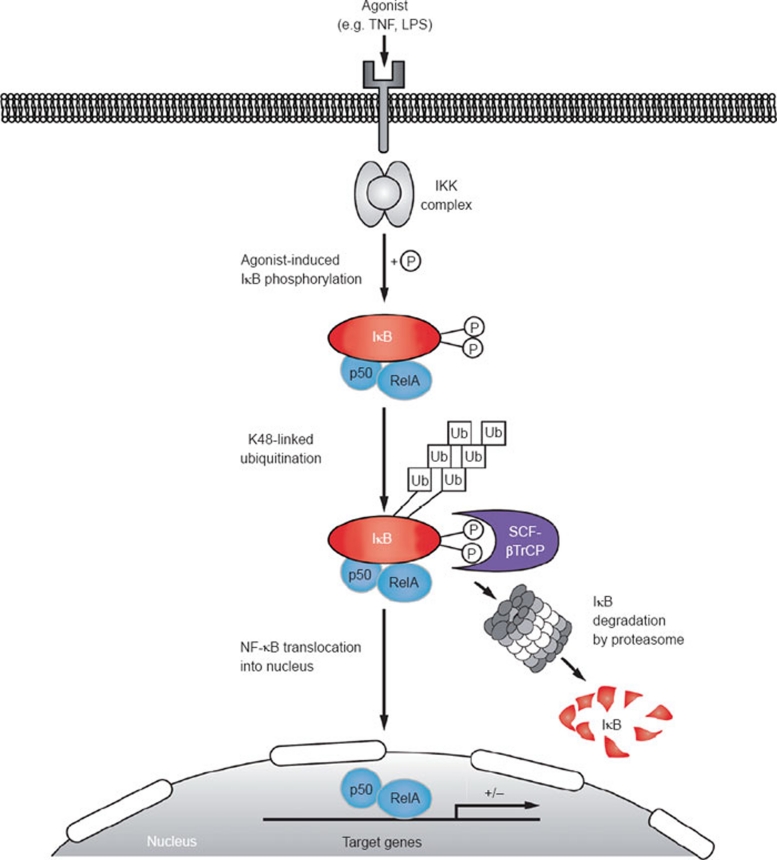
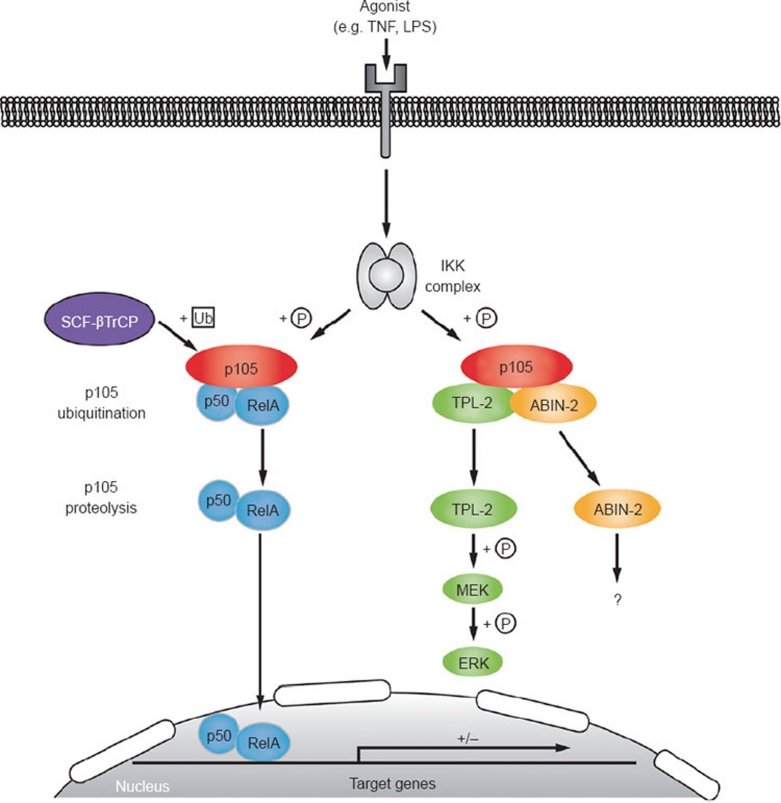
The IκB kinase (IKK) complex plays a well-documented role in innate and adaptive immunity. This function has been widely attributed to its role as the central activator of the NF-κB family of transcription factors. However, another important consequence of IKK activation is the regulation of TPL-2, a MEK kinase that is required for activation of ERK-1/2 MAP kinases in myeloid cells following Toll-like receptor and TNF receptor stimulation. In unstimulated cells, TPL-2 is stoichiometrically complexed with the NF-κB inhibitory protein NF-κB1 p105, which blocks TPL-2 access to its substrate MEK, and the ubiquitin-binding protein ABIN-2 (A20-binding inhibitor of NF-κB 2), both of which are required to maintain TPL-2 protein stability. Following agonist stimulation, the IKK complex phosphorylates p105, triggering its K48-linked ubiquitination and degradation by the proteasome. This releases TPL-2 from p105-mediated inhibition, facilitating activation of MEK, in addition to modulating NF-κB activation by liberating associated Rel subunits for translocation into the nucleus. IKK-induced proteolysis of p105, therefore, can directly regulate both NF-κB and ERK MAP kinase activation via NF-κB1 p105. TPL-2 is critical for production of the proinflammatory cytokine TNF during inflammatory responses. Consequently, there has been considerable interest in the pharmaceutical industry to develop selective TPL-2 inhibitors as drugs for the treatment of TNF-dependent inflammatory diseases, such as rheumatoid arthritis and inflammatory bowel disease. This review summarizes our current understanding of the regulation of TPL-2 signaling function, and also the complex positive and negative roles of TPL-2 in immune and inflammatory responses.
Figures
References
-
- Lund AH, Turner G, Trubetskoy A, et al. Genome-wide retroviral insertional tagging of genes involved in cancer in Cdkn2a-deficient mice. Nat Genet. 2002;32:160–165. - PubMed
-
- Mikkers H, Allen J, Knipscheer P, et al. High throughput retroviral tagging to identify components of specific signaling pathways in cancer. Nat Genet. 2002;32:153–159. - PubMed
-
- Erny KM, Peli J, Lambert JF, Muller V, Diggelmann H. Involvement of the TPL-2/COT oncogene in MMTV tumorigenesis. Oncogene. 1996;13:2015–2020. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
