

Directed adenovirus evolution using engineered mutator viral polymerases
- PMID: 21138963
- PMCID: PMC3061072
- DOI: 10.1093/nar/gkq1258
Directed adenovirus evolution using engineered mutator viral polymerases
Abstract
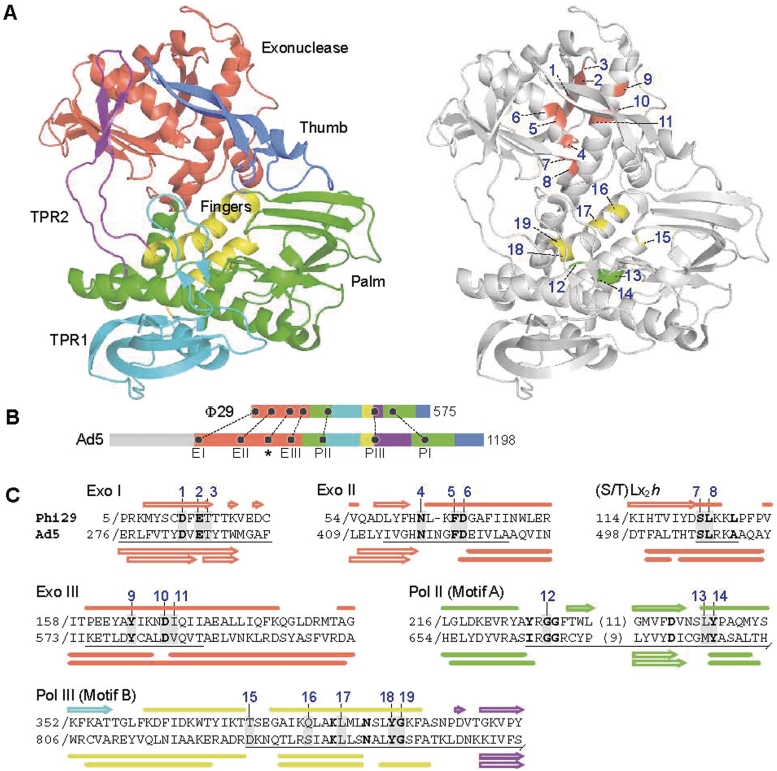
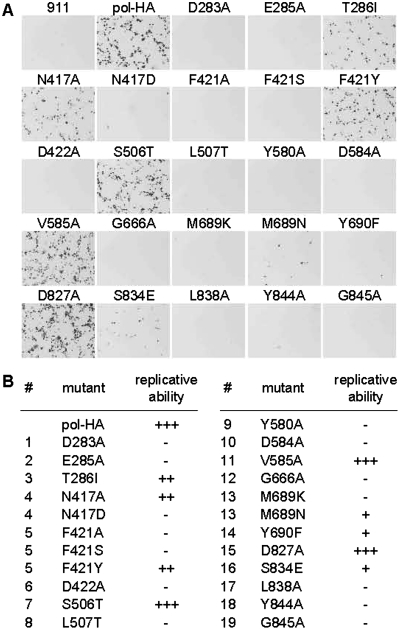
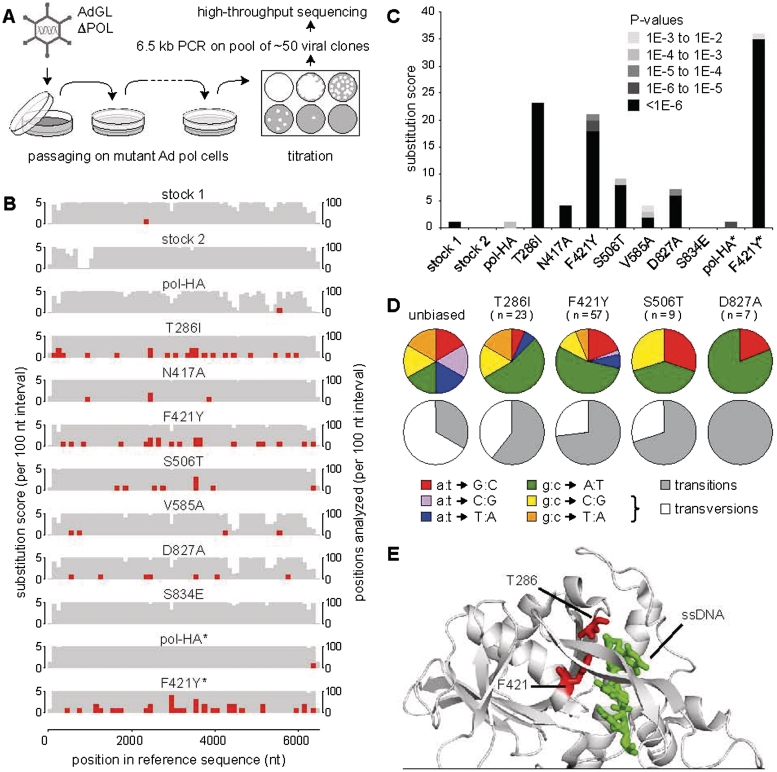
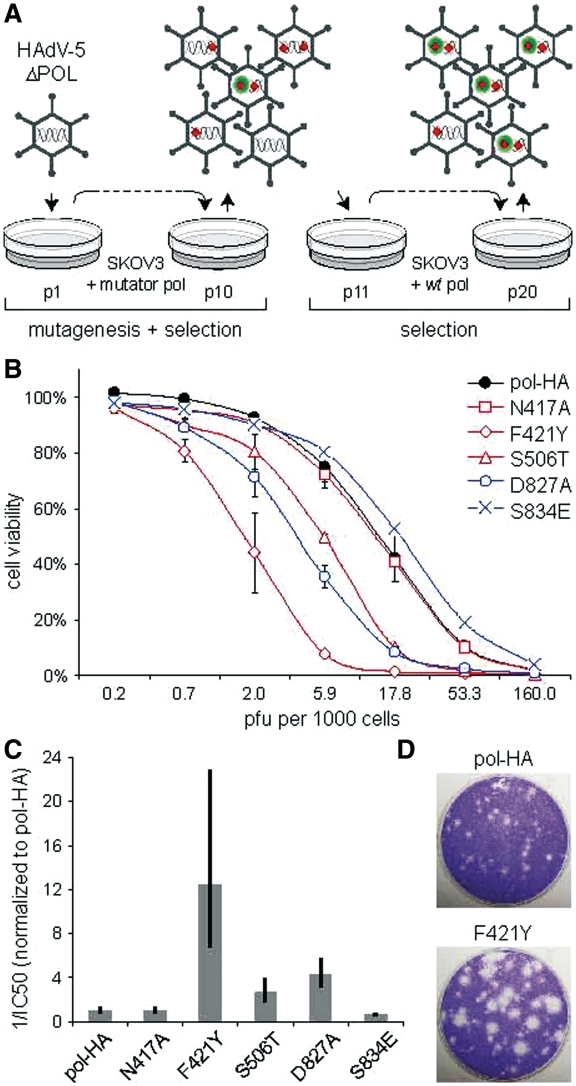
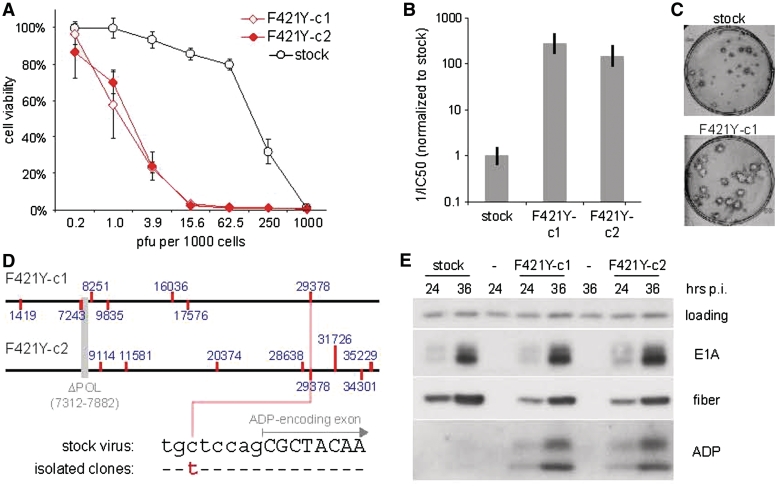
Adenoviruses (Ads) are the most frequently used viruses for oncolytic and gene therapy purposes. Most Ad-based vectors have been generated through rational design. Although this led to significant vector improvements, it is often hampered by an insufficient understanding of Ad's intricate functions and interactions. Here, to evade this issue, we adopted a novel, mutator Ad polymerase-based, 'accelerated-evolution' approach that can serve as general method to generate or optimize adenoviral vectors. First, we site specifically substituted Ad polymerase residues located in either the nucleotide binding pocket or the exonuclease domain. This yielded several polymerase mutants that, while fully supportive of viral replication, increased Ad's intrinsic mutation rate. Mutator activities of these mutants were revealed by performing deep sequencing on pools of replicated viruses. The strongest identified mutators carried replacements of residues implicated in ssDNA binding at the exonuclease active site. Next, we exploited these mutators to generate the genetic diversity required for directed Ad evolution. Using this new forward genetics approach, we isolated viral mutants with improved cytolytic activity. These mutants revealed a common mutation in a splice acceptor site preceding the gene for the adenovirus death protein (ADP). Accordingly, the isolated viruses showed high and untimely expression of ADP, correlating with a severe deregulation of E3 transcript splicing.
Figures

Similar articles
-
Comparison of the E3 and L3 regions for arming oncolytic adenoviruses to achieve a high level of tumor-specific transgene expression.Cancer Gene Ther. 2008 Jan;15(1):9-17. doi: 10.1038/sj.cgt.7701093. Epub 2007 Sep 14. Cancer Gene Ther. 2008. PMID: 17853920
-
ADP-overexpressing adenovirus elicits enhanced cytopathic effect by induction of apoptosis.Cancer Gene Ther. 2005 Jan;12(1):61-71. doi: 10.1038/sj.cgt.7700769. Cancer Gene Ther. 2005. PMID: 15375379
-
Imaging of viral thymidine kinase gene expression by replicating oncolytic adenovirus and prediction of therapeutic efficacy.Yonsei Med J. 2008 Oct 31;49(5):811-8. doi: 10.3349/ymj.2008.49.5.811. Yonsei Med J. 2008. PMID: 18972602 Free PMC article.
-
A compendium of adenovirus genetic modifications for enhanced replication, oncolysis, and tumor immunosurveillance in cancer therapy.Gene. 2018 Dec 30;679:11-18. doi: 10.1016/j.gene.2018.08.069. Epub 2018 Aug 29. Gene. 2018. PMID: 30171937 Review.
-
Non-Human Primate-Derived Adenoviruses for Future Use as Oncolytic Agents?Int J Mol Sci. 2020 Jul 8;21(14):4821. doi: 10.3390/ijms21144821. Int J Mol Sci. 2020. PMID: 32650405 Free PMC article. Review.
Cited by
-
An Adaptable Platform for Directed Evolution in Human Cells.J Am Chem Soc. 2018 Dec 26;140(51):18093-18103. doi: 10.1021/jacs.8b10937. Epub 2018 Dec 14. J Am Chem Soc. 2018. PMID: 30427676 Free PMC article.
-
Current Experimental Studies of Gene Therapy in Parkinson's Disease.Front Aging Neurosci. 2017 May 3;9:126. doi: 10.3389/fnagi.2017.00126. eCollection 2017. Front Aging Neurosci. 2017. PMID: 28515689 Free PMC article. Review.
-
Directed evolution in mammalian cells.Nat Methods. 2021 Apr;18(4):346-357. doi: 10.1038/s41592-021-01090-x. Epub 2021 Apr 7. Nat Methods. 2021. PMID: 33828274 Free PMC article. Review.
-
Combining CRISPR-Cas-mediated terminal resolution with a novel genetic workflow to achieve high-diversity adenoviral libraries.Mol Ther Methods Clin Dev. 2024 Mar 18;32(2):101241. doi: 10.1016/j.omtm.2024.101241. eCollection 2024 Jun 13. Mol Ther Methods Clin Dev. 2024. PMID: 38585687 Free PMC article.
-
Expanding the Scope of Adenoviral Vectors by Utilizing Novel Tools for Recombination and Vector Rescue.Viruses. 2024 Apr 23;16(5):658. doi: 10.3390/v16050658. Viruses. 2024. PMID: 38793540 Free PMC article.
References
-
- Parato KA, Senger D, Forsyth PA, Bell JC. Recent progress in the battle between oncolytic viruses and tumours. Nat. Rev. Cancer. 2005;5:965–976. - PubMed
-
- Alemany R. Designing adenoviral vectors for tumor-specific targeting. Methods Mol. Biol. 2009;542:57–74. - PubMed
-
- Toth K, Dhar D, Wold WS. Oncolytic (replication-competent) adenoviruses as anticancer agents. Expert. Opin. Biol. Ther. 2010;10:353–368. - PubMed
-
- DeWeese TL, van der PH, Li S, Mikhak B, Drew R, Goemann M, Hamper U, DeJong R, Detorie N, Rodriguez R, et al. A phase I trial of CV706, a replication-competent, PSA selective oncolytic adenovirus, for the treatment of locally recurrent prostate cancer following radiation therapy. Cancer Res. 2001;61:7464–7472. - PubMed
-
- Khuri FR, Nemunaitis J, Ganly I, Arseneau J, Tannock IF, Romel L, Gore M, Ironside J, MacDougall RH, Heise C, et al. a controlled trial of intratumoral ONYX-015, a selectively-replicating adenovirus, in combination with cisplatin and 5-fluorouracil in patients with recurrent head and neck cancer. Nat. Med. 2000;6:879–885. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
