

Neuronal plasticity and seasonal reproduction in sheep
- PMID: 21143669
- PMCID: PMC3059776
- DOI: 10.1111/j.1460-9568.2010.07530.x
Neuronal plasticity and seasonal reproduction in sheep
Abstract
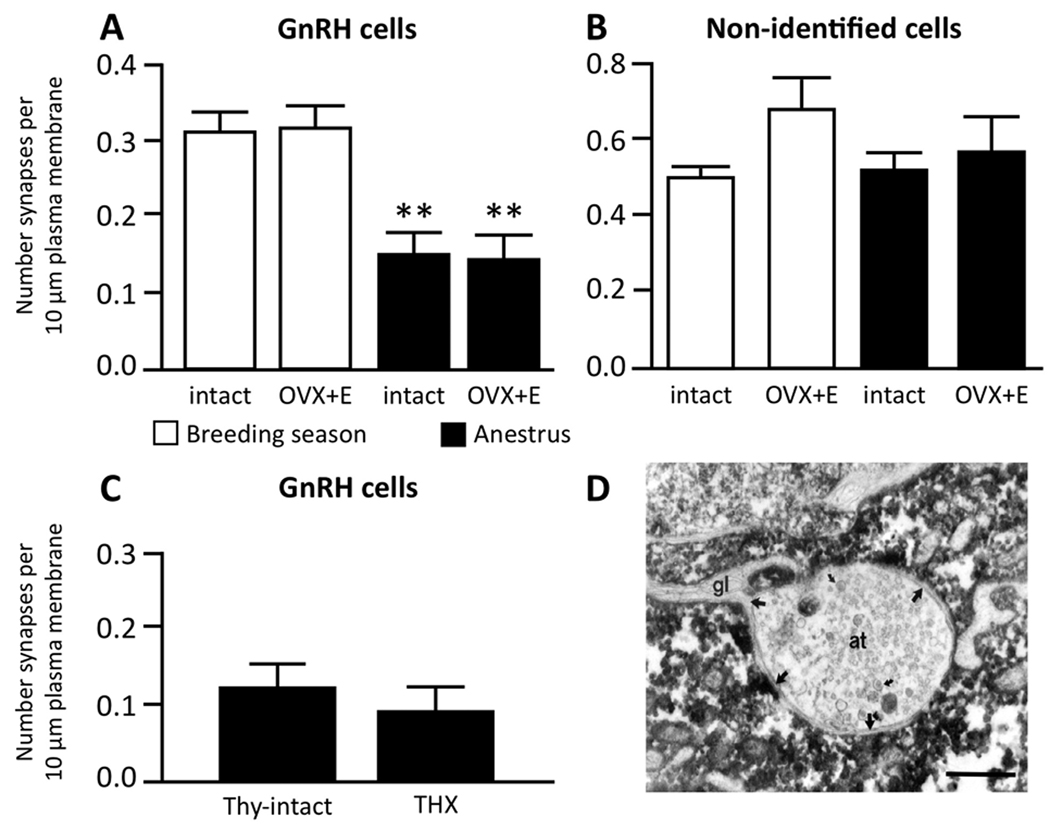
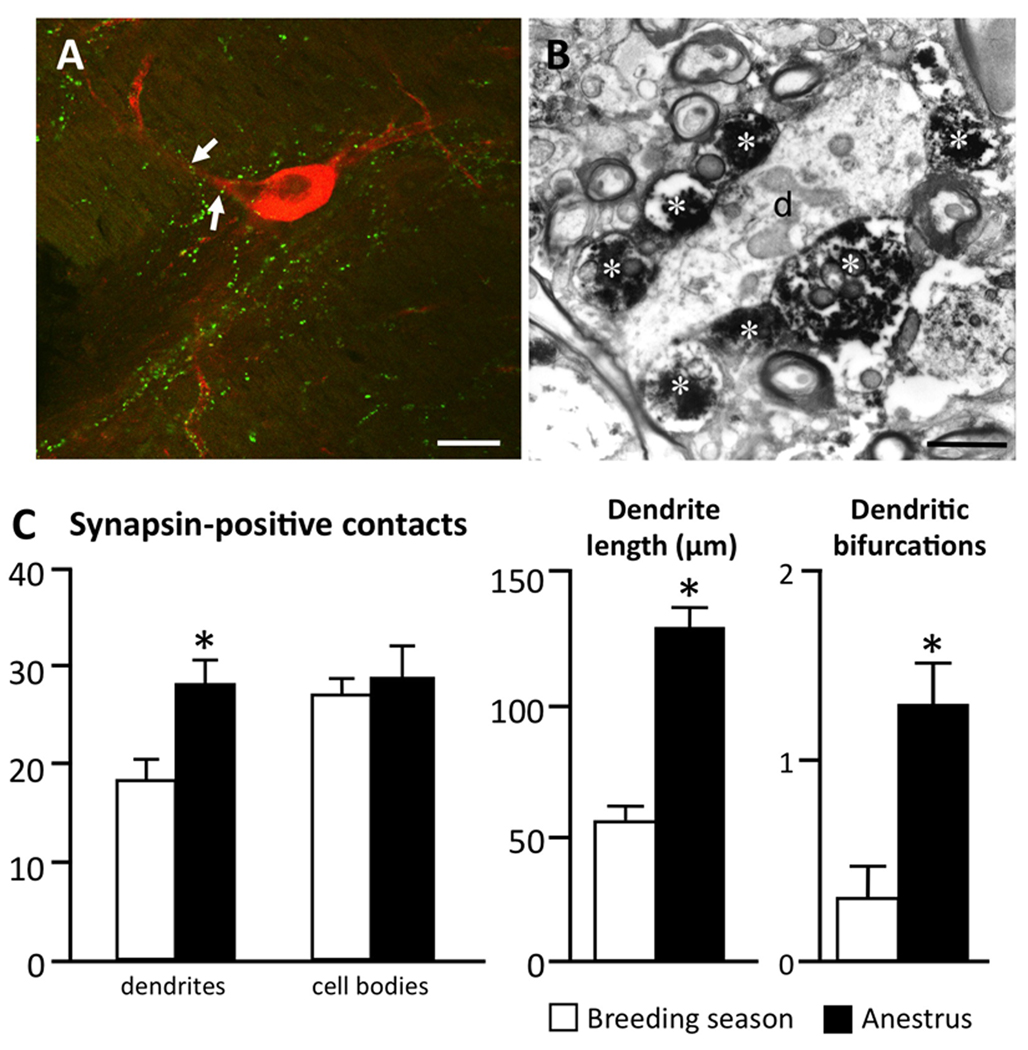
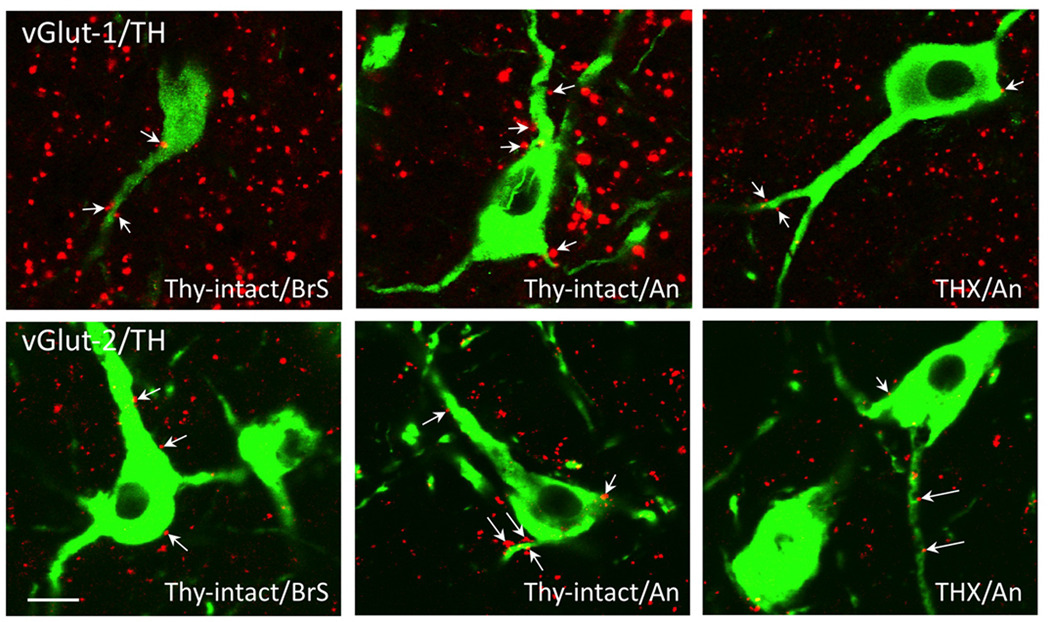
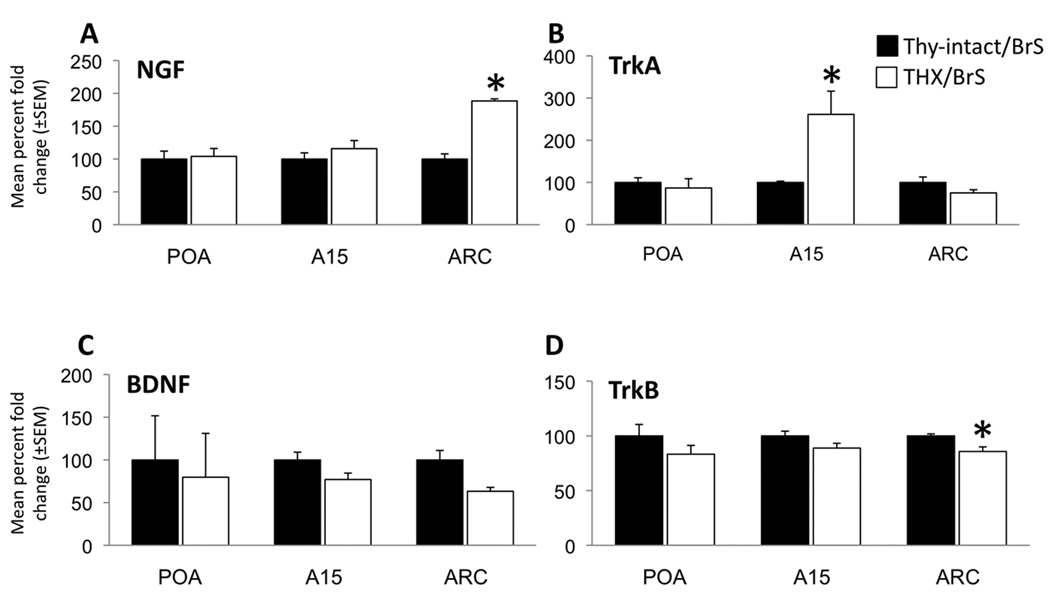
Seasonal reproduction represents a naturally occurring example of functional plasticity in the adult brain as it reflects changes in neuroendocrine pathways controlling gonadotropin-releasing hormone (GnRH) secretion and, in particular, the responsiveness of GnRH neurons to estradiol negative feedback. Structural plasticity within this neural circuitry may, in part, be responsible for seasonal switches in the negative feedback control of GnRH secretion that underlie annual reproductive transitions. We review evidence for structural changes in the circuitry responsible for seasonal inhibition of GnRH secretion in sheep. These include changes in synaptic inputs onto GnRH neurons, as well as onto dopamine neurons in the A15 cell group, a nucleus that plays a key role in estradiol negative feedback. We also present preliminary data suggesting a role for neurotrophins and neurotrophin receptors as an early mechanistic step in the plasticity that accompanies seasonal reproductive transitions in sheep. Finally, we review recent evidence suggesting that kisspeptin cells of the arcuate nucleus constitute a critical intermediary in the control of seasonal reproduction. Although a majority of the data for a role of neuronal plasticity in seasonal reproduction has come from the sheep model, the players and principles are likely to have relevance for reproduction in a wide variety of vertebrates, including humans, and in both health and disease.
© 2010 The Authors. European Journal of Neuroscience © 2010 Federation of European Neuroscience Societies and Blackwell Publishing Ltd.
Figures

Similar articles
-
The GnRH system of seasonal breeders: anatomy and plasticity.Brain Res Bull. 1997;44(4):445-57. doi: 10.1016/s0361-9230(97)00225-6. Brain Res Bull. 1997. PMID: 9370210 Review.
-
Variation in kisspeptin and RFamide-related peptide (RFRP) expression and terminal connections to gonadotropin-releasing hormone neurons in the brain: a novel medium for seasonal breeding in the sheep.Endocrinology. 2008 Nov;149(11):5770-82. doi: 10.1210/en.2008-0581. Epub 2008 Jul 10. Endocrinology. 2008. PMID: 18617612 Free PMC article.
-
Neuroanatomical plasticity in the gonadotropin-releasing hormone system of the ewe: seasonal variation in glutamatergic and gamma-aminobutyric acidergic afferents.J Comp Neurol. 2009 Aug 20;515(6):615-28. doi: 10.1002/cne.22087. J Comp Neurol. 2009. PMID: 19496167
-
Morphological plasticity in the neural circuitry responsible for seasonal breeding in the ewe.Endocrinology. 2006 Oct;147(10):4843-51. doi: 10.1210/en.2006-0408. Epub 2006 Jul 20. Endocrinology. 2006. PMID: 16857749
-
Neural mechanisms controlling seasonal reproduction: principles derived from the sheep model and its comparison with hamsters.Front Neuroendocrinol. 2015 Apr;37:43-51. doi: 10.1016/j.yfrne.2014.12.002. Epub 2015 Jan 9. Front Neuroendocrinol. 2015. PMID: 25582913 Free PMC article. Review.
Cited by
-
Genome-Wide Association Study Reveals Candidate Genes for Litter Size Traits in Pelibuey Sheep.Animals (Basel). 2020 Mar 4;10(3):434. doi: 10.3390/ani10030434. Animals (Basel). 2020. PMID: 32143402 Free PMC article.
-
Effect of immunization against OPN5 on the reproductive performance in Shan Partridge ducks under different photoperiods.Poult Sci. 2024 Apr;103(4):103413. doi: 10.1016/j.psj.2023.103413. Epub 2024 Feb 6. Poult Sci. 2024. PMID: 38442558 Free PMC article.
-
Goats as Valuable Animal Model to Test the Targeted Glutamate Supplementation upon Antral Follicle Number, Ovulation Rate, and LH-Pulsatility.Biology (Basel). 2022 Jul 6;11(7):1015. doi: 10.3390/biology11071015. Biology (Basel). 2022. PMID: 36101396 Free PMC article.
-
Research resource: Gene profiling of G protein-coupled receptors in the arcuate nucleus of the female.Mol Endocrinol. 2014 Aug;28(8):1362-80. doi: 10.1210/me.2014-1103. Epub 2014 Jun 16. Mol Endocrinol. 2014. PMID: 24933249 Free PMC article.
-
Effects of Season and Estradiol on KNDy Neuron Peptides, Colocalization With D2 Dopamine Receptors, and Dopaminergic Inputs in the Ewe.Endocrinology. 2017 Apr 1;158(4):831-841. doi: 10.1210/en.2016-1830. Endocrinology. 2017. PMID: 28324006 Free PMC article.
References
-
- Adams VL, Goodman RL, Salm AK, Coolen LM, Karsch FJ, Lehman MN. Morphological plasticity in the neural circuitry responsible for seasonal breeding in the ewe. Endocrinology. 2006;147:4843–4851. - PubMed
-
- Alvarez-Dolado M, Iglesias T, Rodriguez-Pena A, Bernal J, Munoz A. Expression of neurotrophins and the trk family of neurotrophin receptors in normal and hypothyroid rat brain. Brain Res Mol Brain Res. 1994;27:249–257. - PubMed
-
- Anderson GM, Barrell GK. Pulsatile luteinizing hormone secretion in the ovariectomized, thyroidectomized red deer hind following treatment with dopaminergic and opioidergic agonists and antagonists. Biol Reprod. 1998;59:960–968. - PubMed
-
- Anderson GM, Connors JM, Hardy SL, Valent M, Goodman RL. Oestradiol microimplants in the ventromedial preoptic area inhibit secretion of luteinizing hormone via dopamine neurones in anoestrous ewes. J Neuroendocrinol. 2001;13:1051–1058. - PubMed
-
- Anderson GM, Hardy SL, Valent M, Billings HJ, Connors JM, Goodman RL. Evidence that thyroid hormones act in the ventromedial preoptic area and the premammillary region of the brain to allow the termination of the breeding season in the ewe. Endocrinology. 2003;144:2892–2901. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
