

Programmed cell death-1 (PD-1) at the heart of heterologous prime-boost vaccines and regulation of CD8+ T cell immunity
- PMID: 21144062
- PMCID: PMC3012026
- DOI: 10.1186/1479-5876-8-132
Programmed cell death-1 (PD-1) at the heart of heterologous prime-boost vaccines and regulation of CD8+ T cell immunity
Abstract
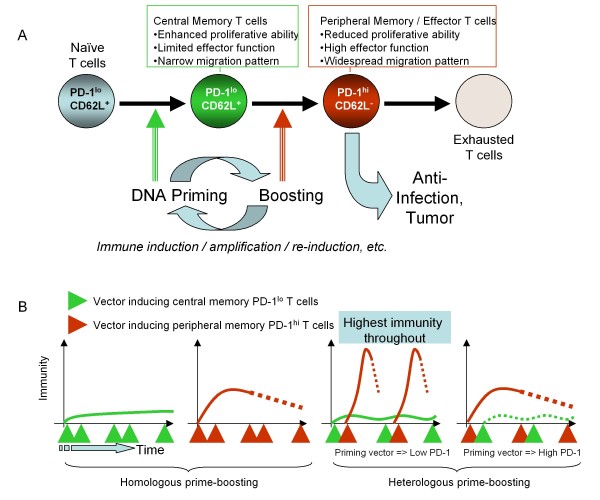
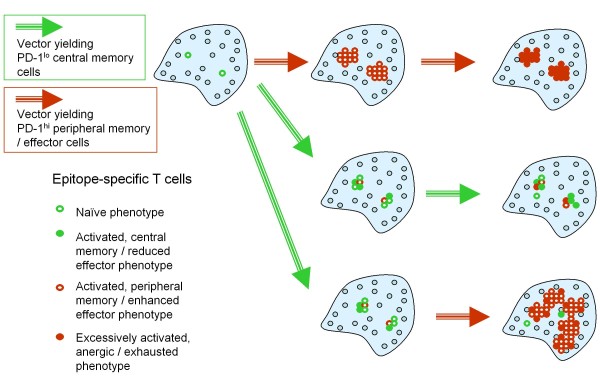
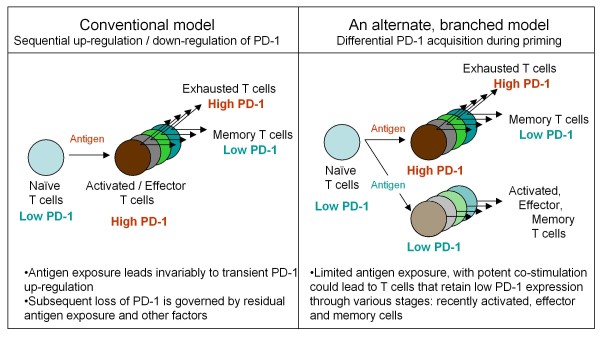
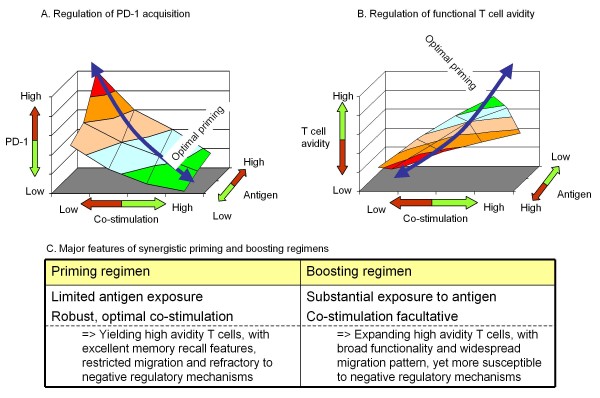
Developing new vaccination strategies and optimizing current vaccines through heterologous prime-boost carries the promise of integrating the benefits of different yet synergistic vectors. It has been widely thought that the increased immunity afforded by heterologous prime-boost vaccination is mainly due to the minimization of immune responses to the carrier vectors, which allows a progressive build up of immunity against defined epitopes and the subsequent induction of broader immune responses against pathogens. Focusing on CD8+ T cells, we put forward a different yet complementary hypothesis based primarily on the systematic analysis of DNA vaccines as priming agents. This hypothesis relies on the finding that during the initiation of immune response, acquisition of co-inhibitory receptors such as programmed cell death-1 (PD-1) is determined by the pattern of antigen exposure in conjunction with Toll-like receptor (TLR)-dependent stimulation, critically affecting the magnitude and profile of secondary immunity. This hypothesis, based upon the acquisition and co-regulation of pivotal inhibitory receptors by CD8+ T cells, offers a rationale for gene-based immunization as an effective priming strategy and, in addition, outlines a new dimension to immune homeostasis during immune reaction to pathogens. Finally, this model implies that new and optimized immunization approaches for cancer and certain viral infections must induce highly efficacious T cells, refractory to a broad range of immune-inhibiting mechanisms, rather than solely or primarily focusing on the generation of large pools of vaccine-specific lymphocytes.
Figures
References
-
- Kenter GG, Welters MJ, Valentijn AR, Lowik MJ, Berends-van der Meer DM, Vloon AP, Essahsah F, Fathers LM, Offringa R, Drijfhout JW, Wafelman AR, Oostendorp J, Fleuren GJ, van der Burg SH, Melief CJ. Vaccination against HPV-16 oncoproteins for vulvar intraepithelial neoplasia. N Engl J Med. 2010;363:943–53. doi: 10.1056/NEJMoa0908806. - DOI - PubMed
-
- Morse MA, Whelan M. A year of successful cancer vaccines points to a path forward. Curr Opin Mol Ther. 2010;12:11–13. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
