

M1 muscarinic receptors boost synaptic potentials and calcium influx in dendritic spines by inhibiting postsynaptic SK channels
- PMID: 21145006
- PMCID: PMC3052967
- DOI: 10.1016/j.neuron.2010.09.004
M1 muscarinic receptors boost synaptic potentials and calcium influx in dendritic spines by inhibiting postsynaptic SK channels
Abstract
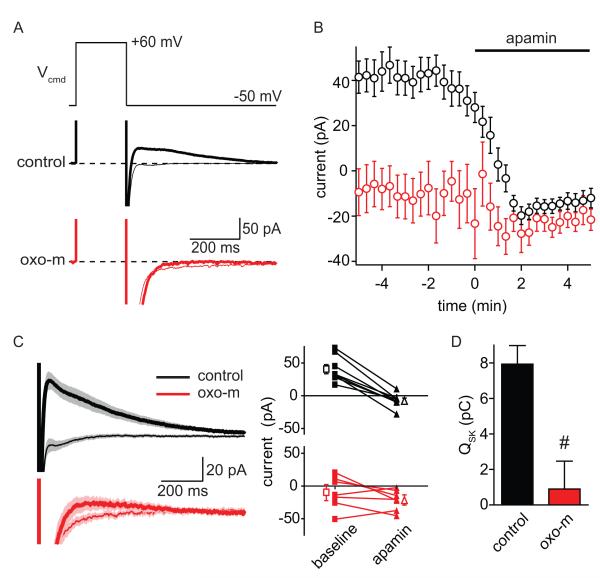
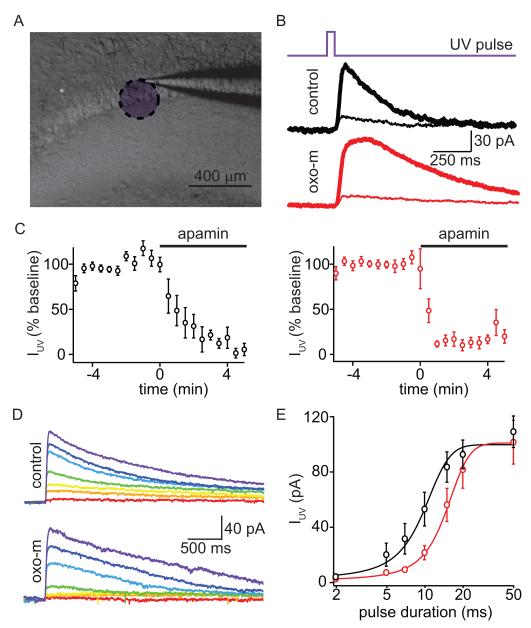
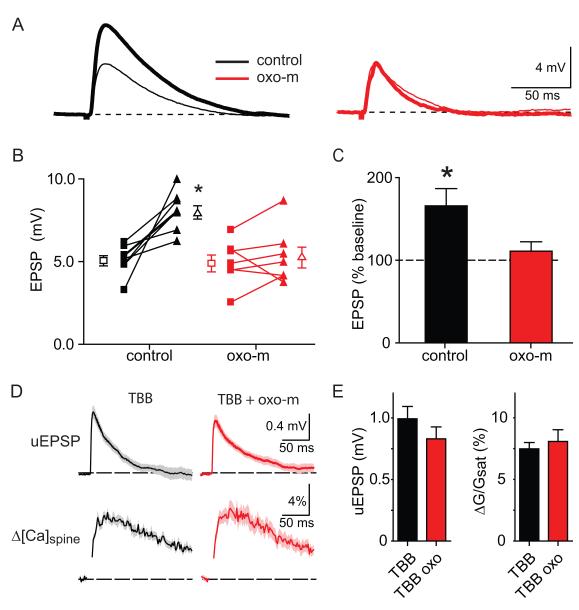
Acetylcholine release and activation of muscarinic cholinergic receptors (mAChRs) enhance synaptic plasticity in vitro and cognition and memory in vivo. Within the hippocampus, mAChRs promote NMDA-type glutamate receptor-dependent forms of long-term potentiation. Here, we use calcium (Ca) imaging combined with two-photon laser glutamate uncaging at apical spines of CA1 pyramidal neurons to examine postsynaptic mechanisms of muscarinic modulation of glutamatergic transmission. Uncaging-evoked excitatory postsynaptic potentials and Ca transients are increased by muscarinic stimulation; however, this is not due to direct modulation of glutamate receptors. Instead, mAChRs modulate a negative feedback loop in spines that normally suppresses synaptic signals. mAChR activation reduces the Ca sensitivity of small conductance Ca-activated potassium (SK) channels that are found in the spine, resulting in increased synaptic potentials and Ca transients. These effects are mediated by M1-type muscarinic receptors and occur in a casein kinase-2-dependent manner. Thus, muscarinic modulation regulates synaptic transmission by tuning the activity of nonglutamatergic postsynaptic ion channels.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
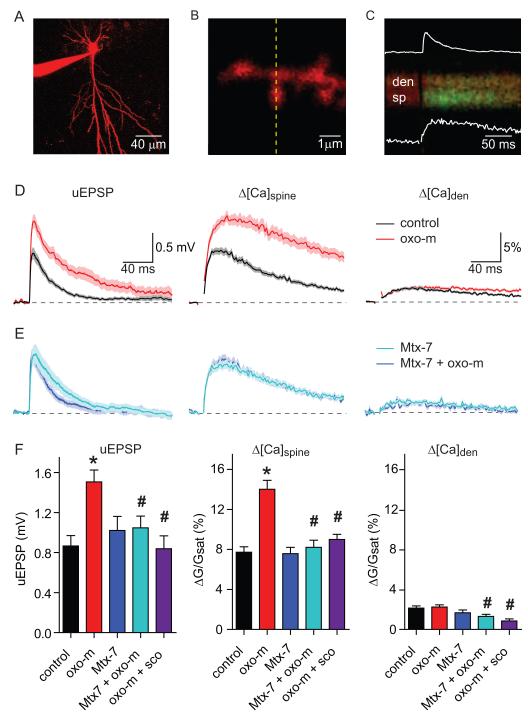
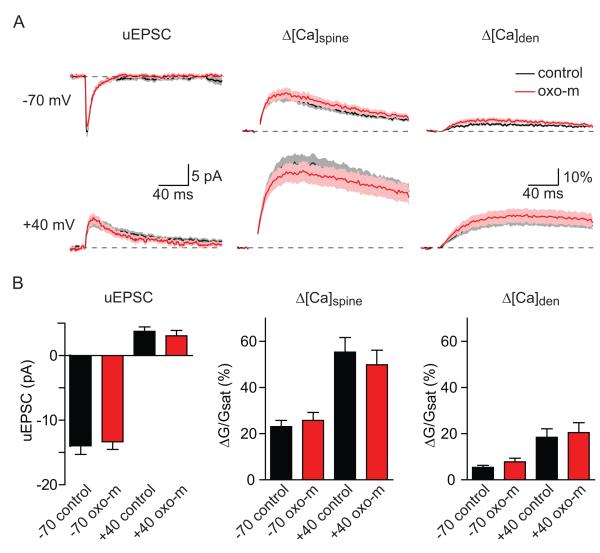
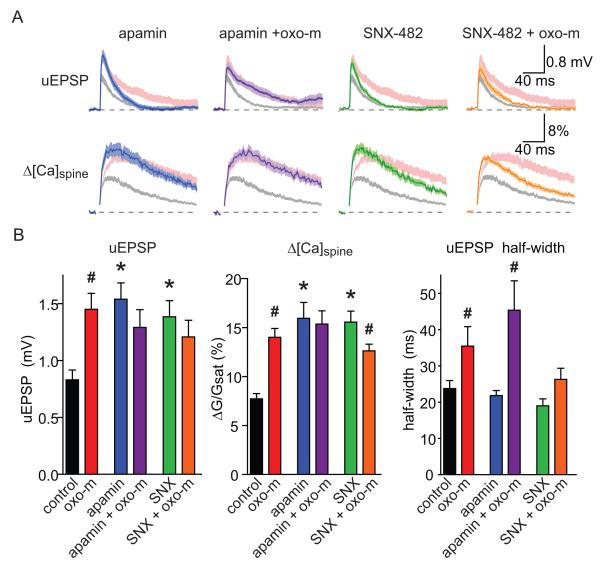
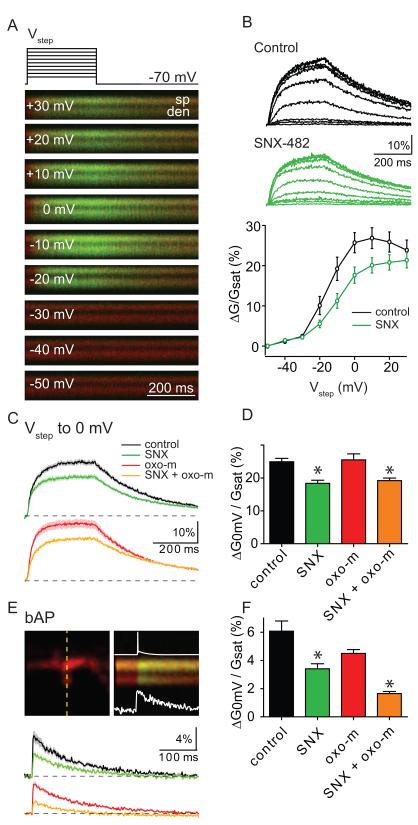
Comment in
-
Cholinergic signaling through synaptic SK channels: it's a protein kinase but which one?Neuron. 2010 Dec 9;68(5):809-11. doi: 10.1016/j.neuron.2010.11.037. Neuron. 2010. PMID: 21144995 Free PMC article.
-
Synaptic plasticity: breaking the learning barrier with ACh.Nat Rev Neurosci. 2011 Feb;12(2):62. doi: 10.1038/nrn2986. Nat Rev Neurosci. 2011. PMID: 21309090 No abstract available.
References
-
- Bildl W, Strassmaier T, Thurm H, Andersen J, Eble S, Oliver D, Knipper M, Mann M, Schulte U, Adelman JP, Fakler B. Protein kinase CK2 is coassembled with small conductance Ca(2+)-activated K+ channels and regulates channel gating. Neuron. 2004;43:847–858. - PubMed
-
- Bloodgood BL, Sabatini BL. Nonlinear regulation of unitary synaptic signals by CaV(2.3) voltage-sensitive calcium channels located in dendritic spines. Neuron. 2007;53:249–260. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
