

Drosophila pacemaker neurons require g protein signaling and GABAergic inputs to generate twenty-four hour behavioral rhythms
- PMID: 21145008
- PMCID: PMC3030199
- DOI: 10.1016/j.neuron.2010.11.017
Drosophila pacemaker neurons require g protein signaling and GABAergic inputs to generate twenty-four hour behavioral rhythms
Abstract
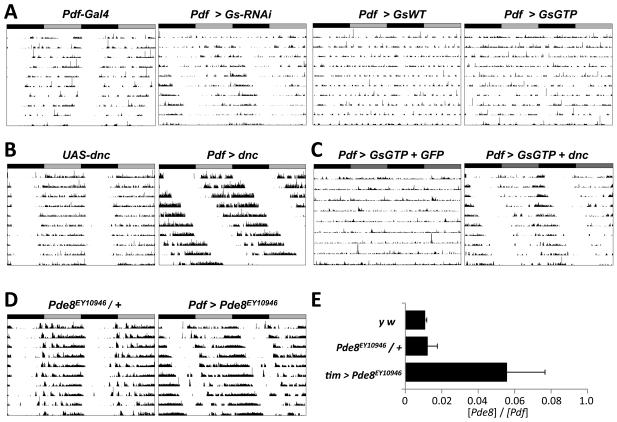
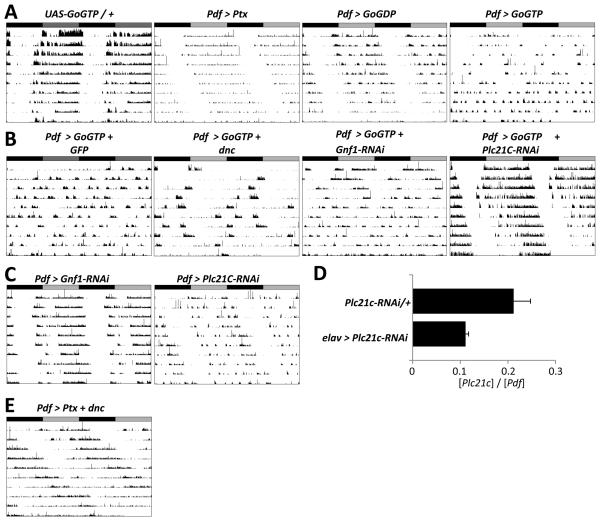
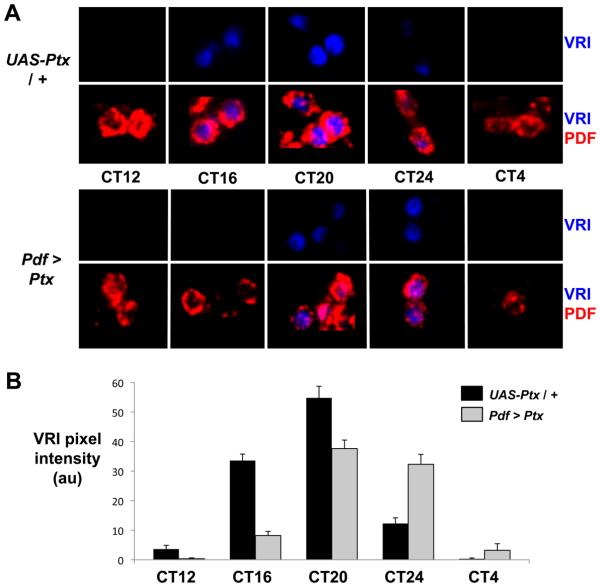
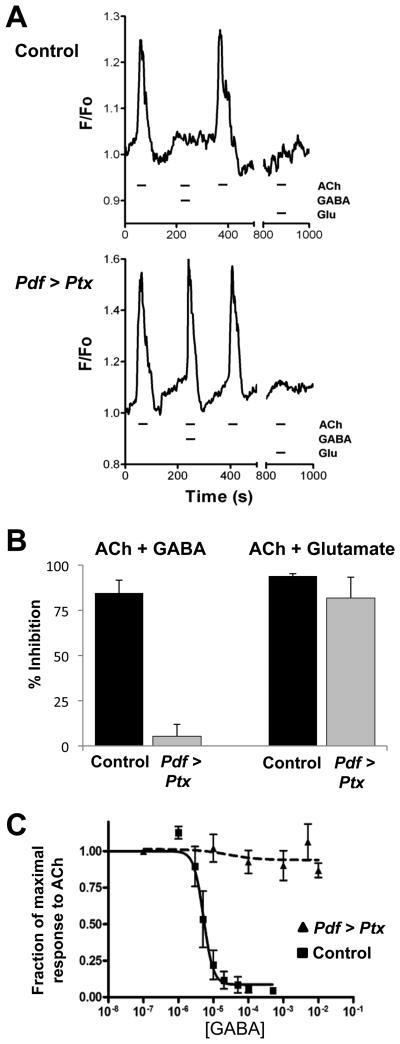
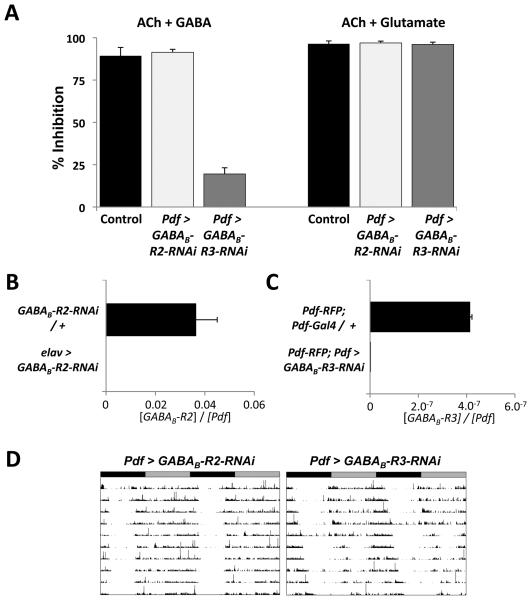
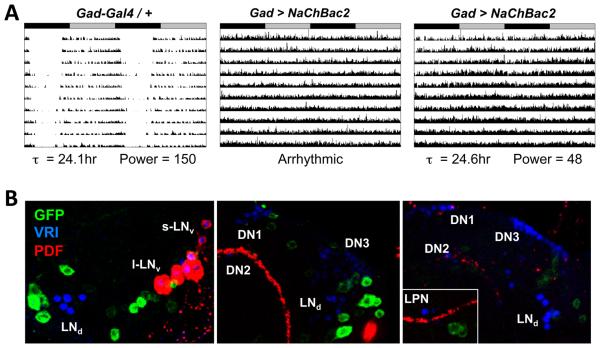
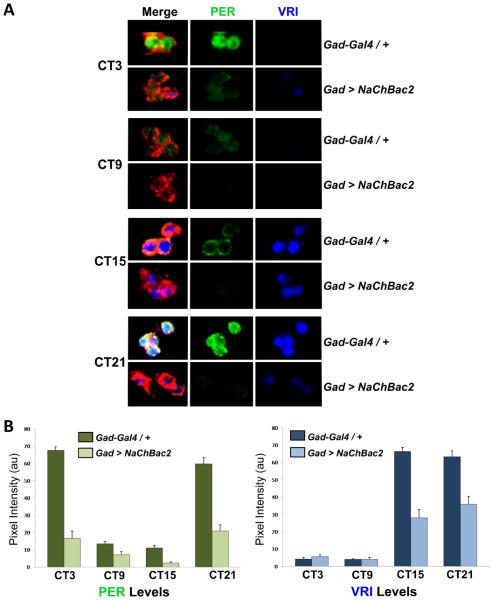
Intercellular signaling is important for accurate circadian rhythms. In Drosophila, the small ventral lateral neurons (s-LN(v)s) are the dominant pacemaker neurons and set the pace of most other clock neurons in constant darkness. Here we show that two distinct G protein signaling pathways are required in LN(v)s for 24 hr rhythms. Reducing signaling in LN(v)s via the G alpha subunit Gs, which signals via cAMP, or via the G alpha subunit Go, which we show signals via Phospholipase 21c, lengthens the period of behavioral rhythms. In contrast, constitutive Gs or Go signaling makes most flies arrhythmic. Using dissociated LN(v)s in culture, we found that Go and the metabotropic GABA(B)-R3 receptor are required for the inhibitory effects of GABA on LN(v)s and that reduced GABA(B)-R3 expression in vivo lengthens period. Although no clock neurons produce GABA, hyperexciting GABAergic neurons disrupts behavioral rhythms and s-LN(v) molecular clocks. Therefore, s-LN(v)s require GABAergic inputs for 24 hr rhythms.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
References
-
- Allada R, Emery P, Takahashi JS, Rosbash M. Stopping time: the genetics of fly and mouse circadian clocks. Annu Rev Neurosci. 2001;24:1091–1119. - PubMed
-
- Balsalobre A, Damiola F, Schibler U. A serum shock induces circadian gene expression in mammalian tissue culture cells. Cell. 1998;93:929–937. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
