

The chromatin signaling pathway: diverse mechanisms of recruitment of histone-modifying enzymes and varied biological outcomes
- PMID: 21145479
- PMCID: PMC3037032
- DOI: 10.1016/j.molcel.2010.11.031
The chromatin signaling pathway: diverse mechanisms of recruitment of histone-modifying enzymes and varied biological outcomes
Abstract
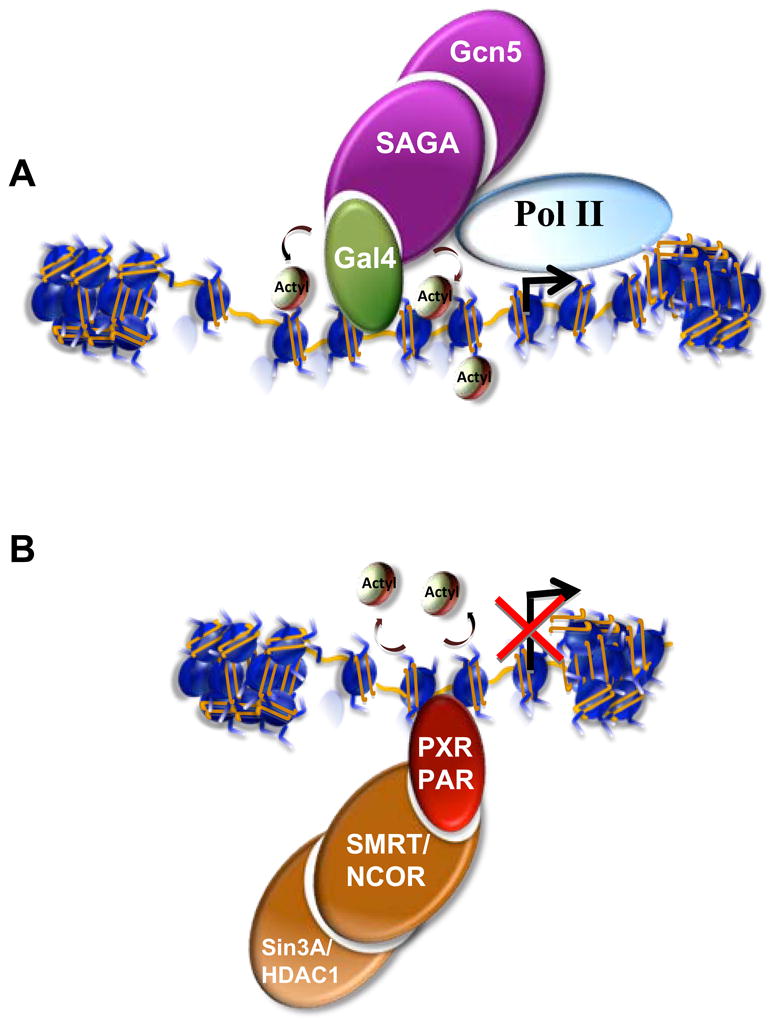
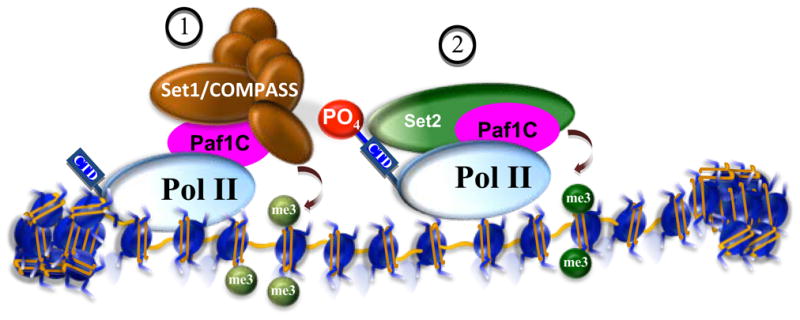
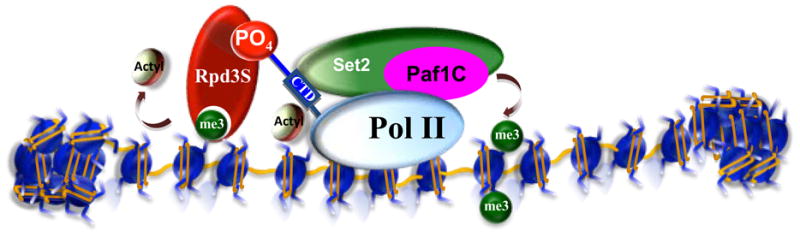
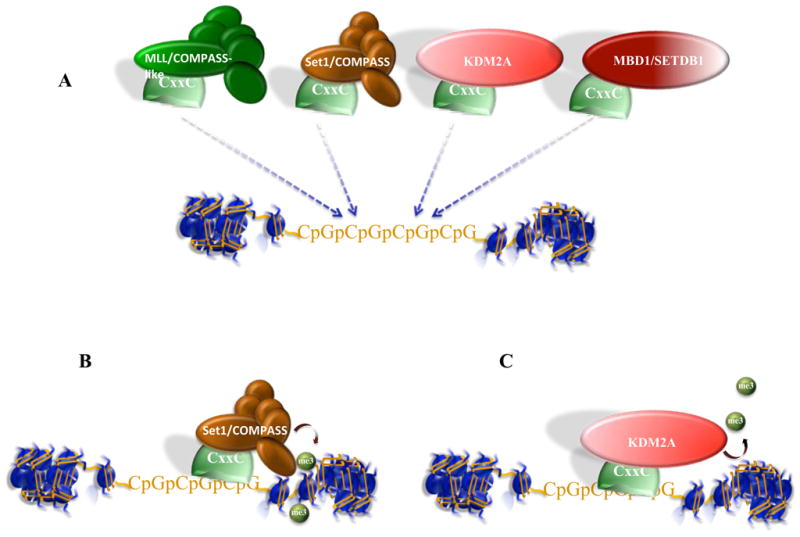
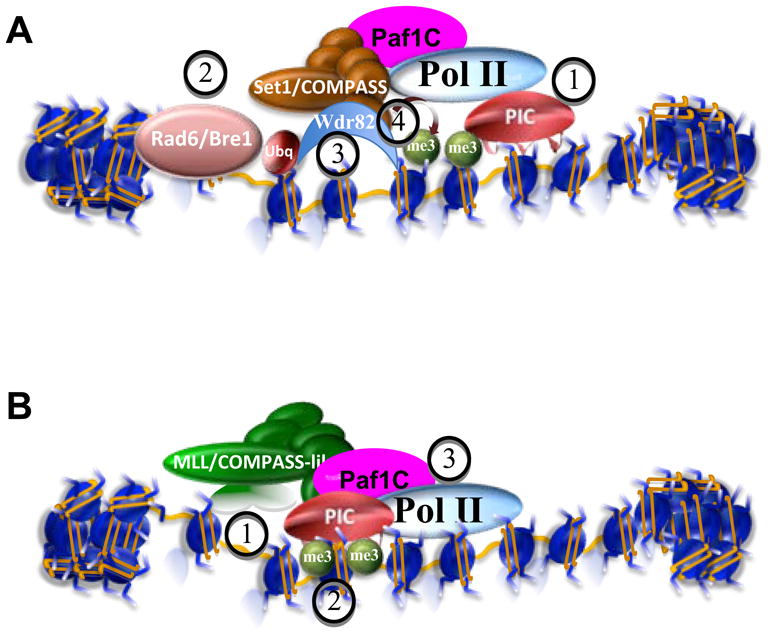
Posttranslational modifications of histones are coupled in the regulation of the cellular processes involving chromatin, such as transcription, replication, repair, and genome stability. Recent biochemical and genetic studies have clearly demonstrated that many aspects of chromatin, in addition to posttranslational modifications of histones, provide surfaces that can interact with effectors and the modifying machineries in a context-dependent manner, all as a part of the "chromatin signaling pathway." Here, we have reviewed recent findings on the molecular basis for the recruitment of the chromatin-modifying machineries and their diverse and varied biological outcomes.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
References
-
- Alland L, Muhle R, Hou H, Jr, Potes J, Chin L, Schreiber-Agus N, DePinho RA. Role for N-CoR and histone deacetylase in Sin3-mediated transcriptional repression. Nature. 1997;387:49–55. - PubMed
-
- Bach C, Mueller D, Buhl S, Garcia-Cuellar MP, Slany RK. Alterations of the CxxC domain preclude oncogenic activation of mixed-lineage leukemia 2. Oncogene. 2009;28:815–823. - PubMed
-
- Bannister AJ, Kouzarides T. The CBP co-activator is a histone acetyltransferase. Nature. 1996;384:641–643. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
