

Bioenergetic function in cardiovascular cells: the importance of the reserve capacity and its biological regulation
- PMID: 21147079
- PMCID: PMC3090710
- DOI: 10.1016/j.cbi.2010.12.002
Bioenergetic function in cardiovascular cells: the importance of the reserve capacity and its biological regulation
Abstract
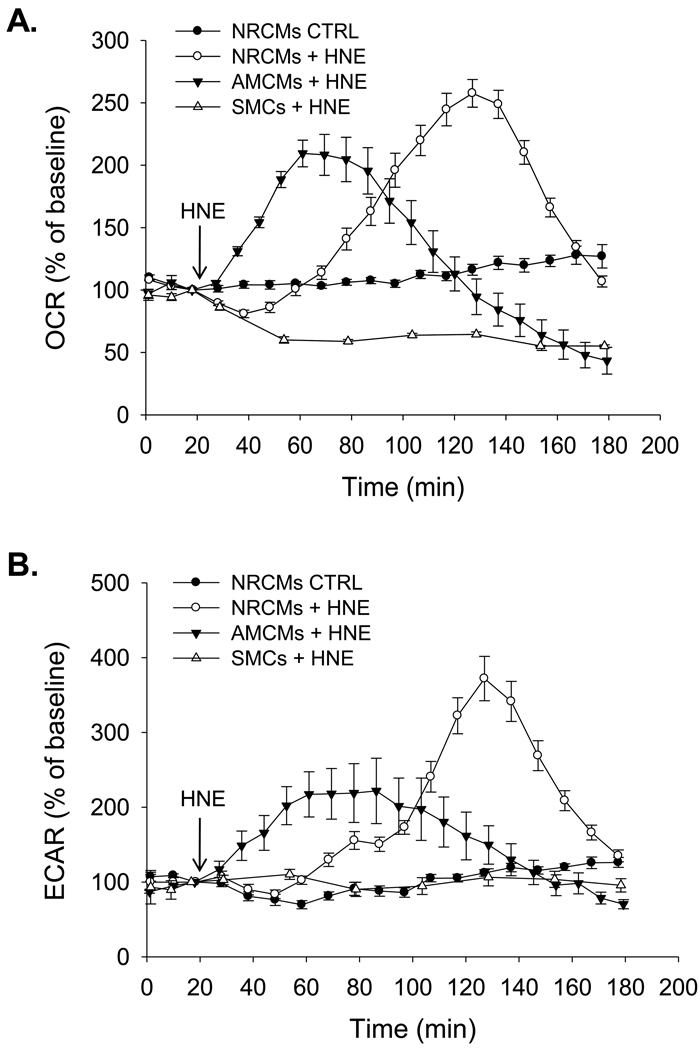
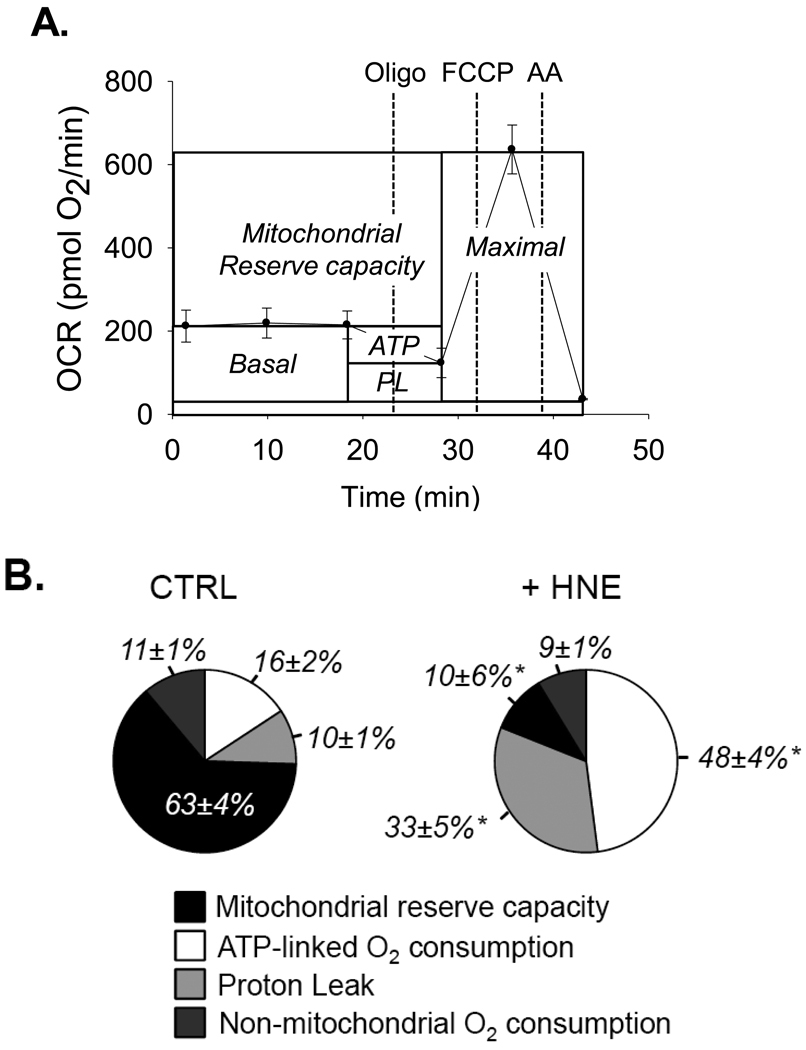
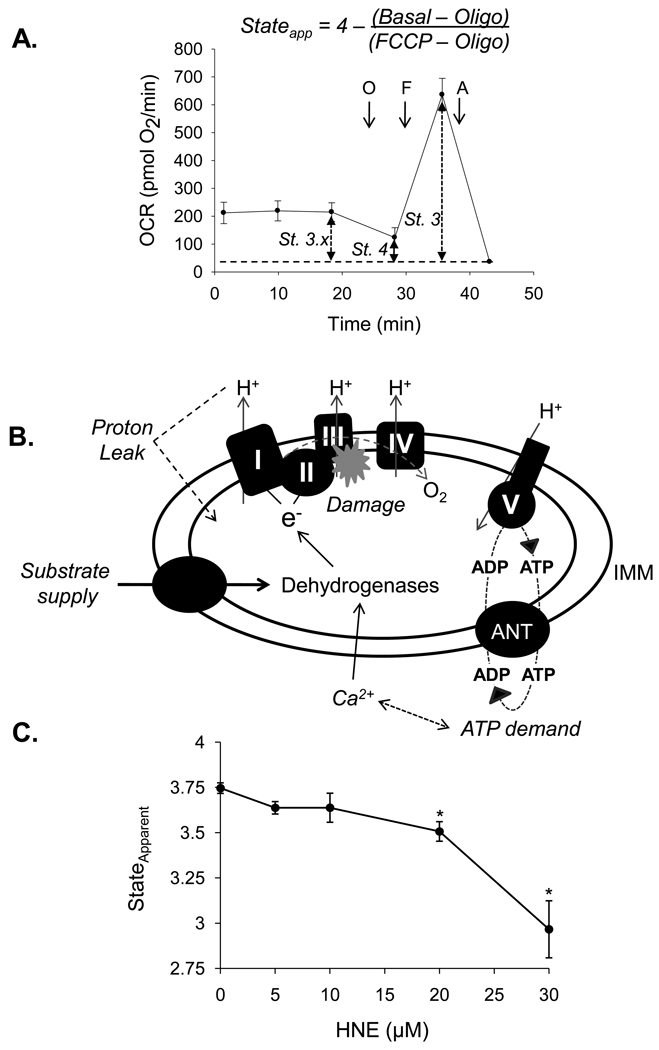
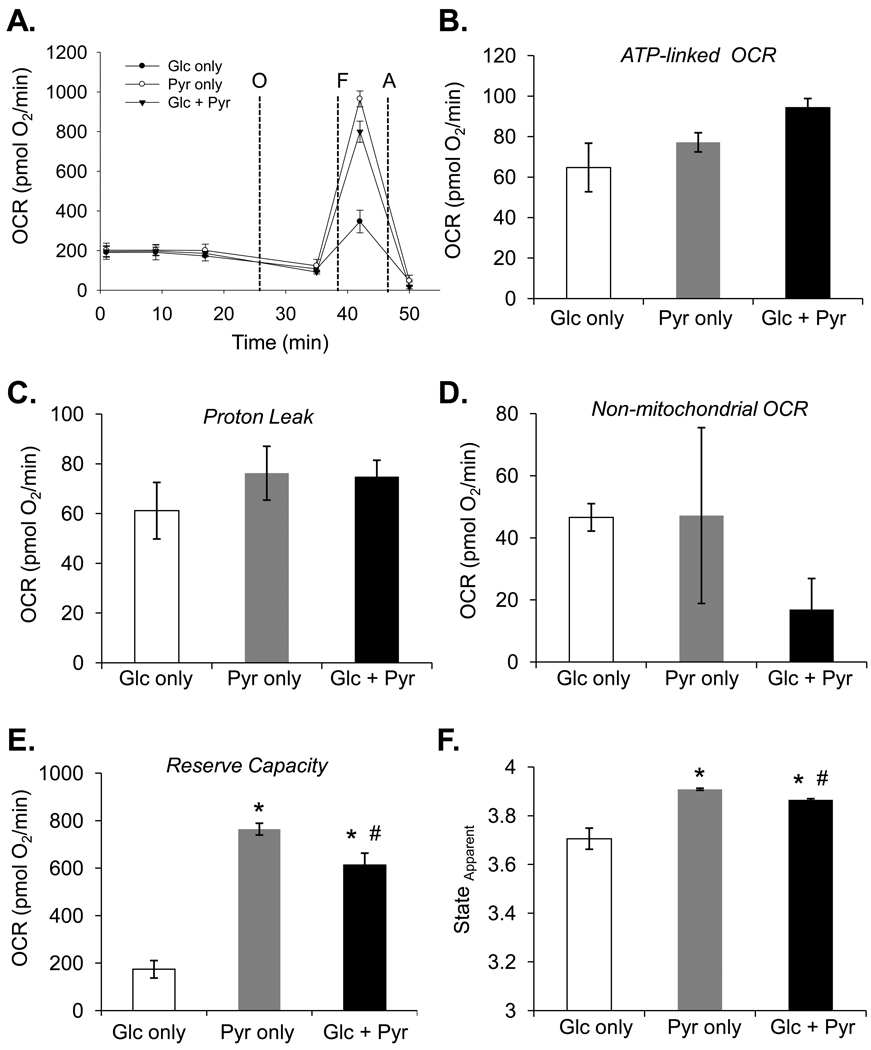
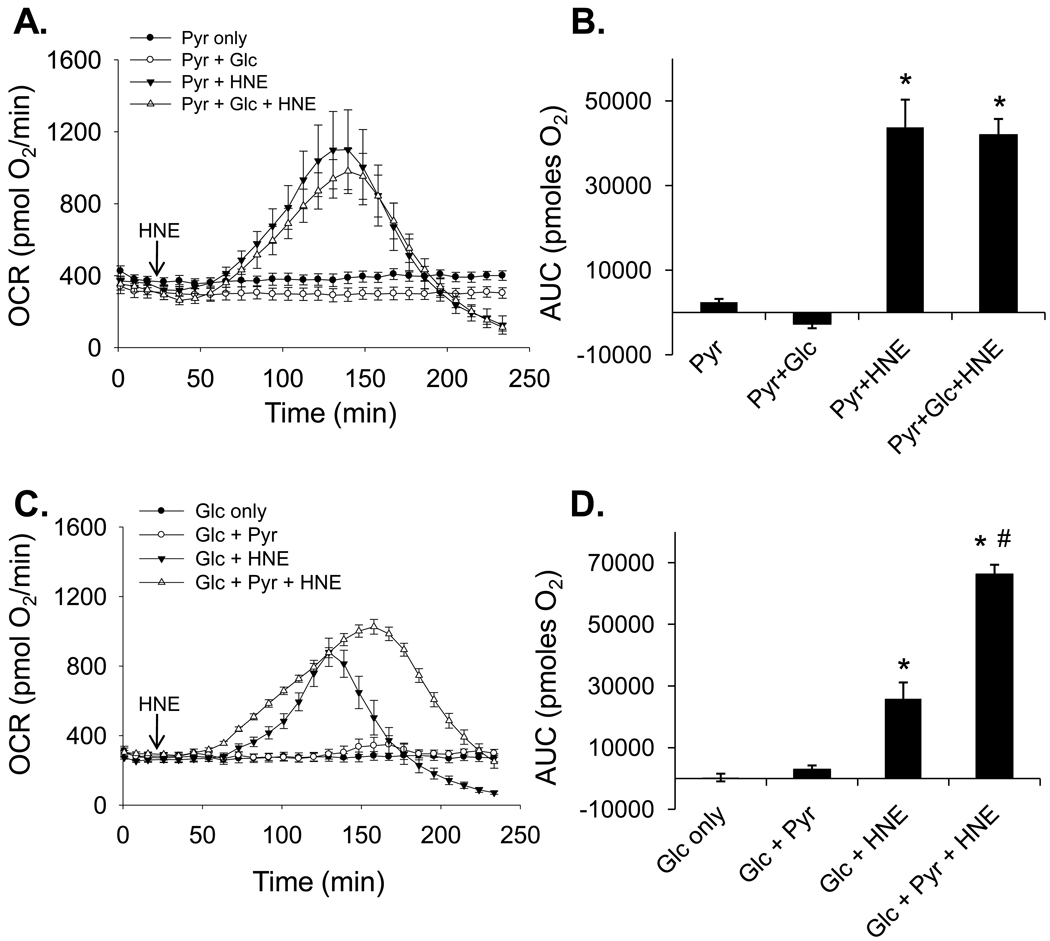
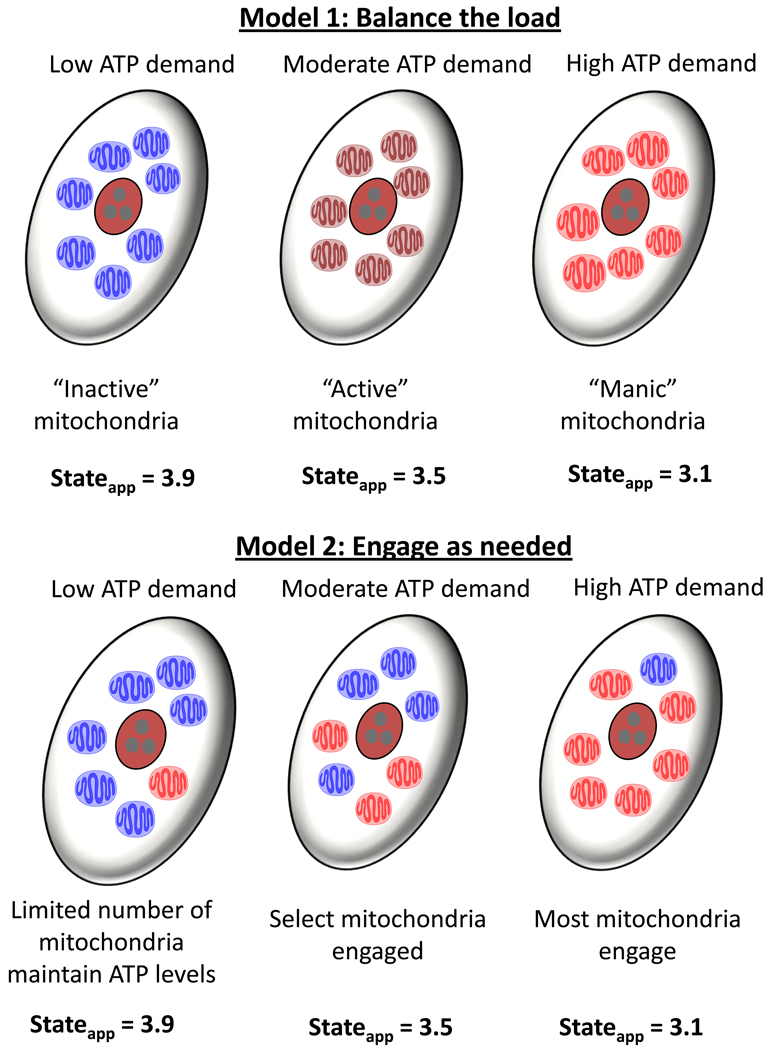
The ability of the cell to generate sufficient energy through oxidative phosphorylation and to maintain healthy pools of mitochondria are critical for survival and maintenance of normal biological function, especially during periods of increased oxidative stress. Mitochondria in most cardiovascular cells function at a basal level that only draws upon a small fraction of the total bioenergetic capability of the organelle; the apparent respiratory state of mitochondria in these cells is often close to state 4. The difference between the basal and maximal activity, equivalent to state 3, of the respiratory chain is called the reserve capacity. We hypothesize that the reserve capacity serves the increased energy demands for maintenance of organ function and cellular repair. However, the factors that determine the volume of the reserve capacity and its relevance to biology are not well understood. In this study, we first examined whether responses to 4-hydroxynonenal (HNE), a lipid peroxidation product found in atherosclerotic lesions and the diseased heart, differ between vascular smooth muscle cells, adult mouse cardiomyocytes, and rat neonatal cardiomyocytes. In both types of cardiomyocytes, oxygen consumption increased after HNE treatment, while oxygen consumption in smooth muscle cells decreased. The increase in oxygen consumption in cardiomyocytes decreased the reserve capacity and shifted the apparent respiratory state closer to state 3. Neonatal rat cardiomyocytes respiring on pyruvate alone had a fourfold higher reserve capacity than cells with glucose as the sole substrate, and these cells were more resistant to mitochondrial dysfunction induced by 4-HNE. The integration of the concepts of reserve capacity and state-apparent are discussed along with the proposal of two potential models by which mitochondria respond to stress.
Copyright © 2010 Elsevier Ireland Ltd. All rights reserved.
Figures
References
-
- Akao M, Ohler A, O'Rourke B, Marban E. Mitochondrial ATP-sensitive potassium channels inhibit apoptosis induced by oxidative stress in cardiac cells. Circ Res. 2001;88:1267–1275. - PubMed
-
- Brand MD. The proton leak across the mitochondrial inner membrane. Biochim Biophys Acta. 1990;1018:128–133. - PubMed
-
- Brown GC. The leaks and slips of bioenergetic membranes. FASEB J. 1992;6:2961–2965. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
