

IgA-producing plasma cells originate from germinal centers that are induced by B-cell receptor engagement in humans
- PMID: 21147106
- PMCID: PMC7115992
- DOI: 10.1053/j.gastro.2010.12.005
IgA-producing plasma cells originate from germinal centers that are induced by B-cell receptor engagement in humans
Abstract
Background & aims: IgA contributes to homeostatic balance between host and intestinal microbiota. Mechanisms that initiate the IgA response are unclear and likely to differ between humans and animal models. We used multiple experimental approaches to investigate the origin of human intestinal plasma cells that produce IgA in the gastrointestinal tract.
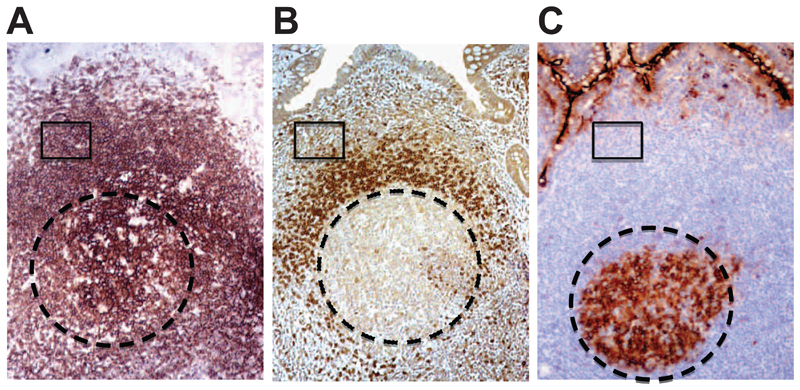
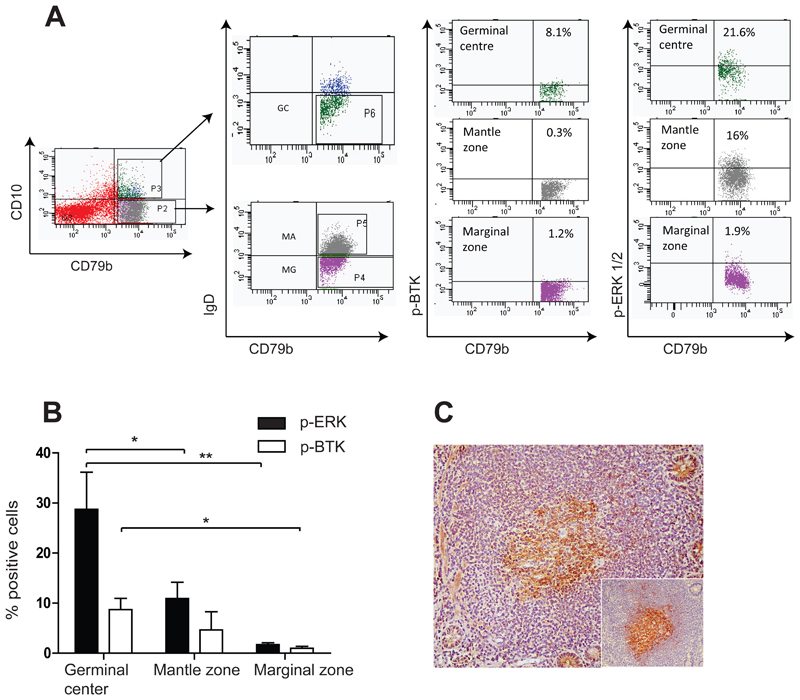
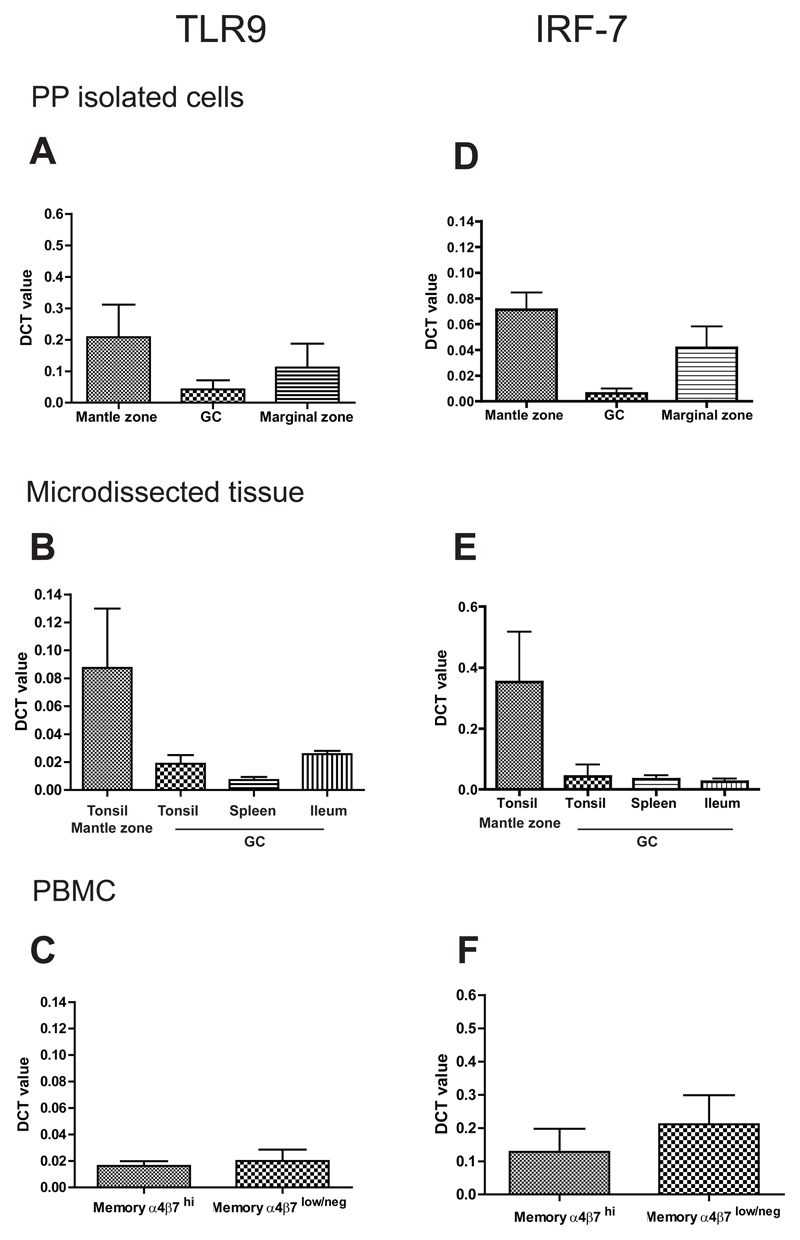
Methods: Complexity of IgA-producing plasma cell populations in human gastrointestinal mucosa and bone marrow and the specific response to oral cholera vaccine were compared by analysis of immunoglobulin genes. Flow cytometry, gene expression analysis, and immunohistochemistry were used to analyze signaling pathways induced by B-cell receptor engagement in human gut-associated lymphoid tissue (GALT) and involvement of innate immunity in B-cell activation in GALT compared with nonintestinal sites.
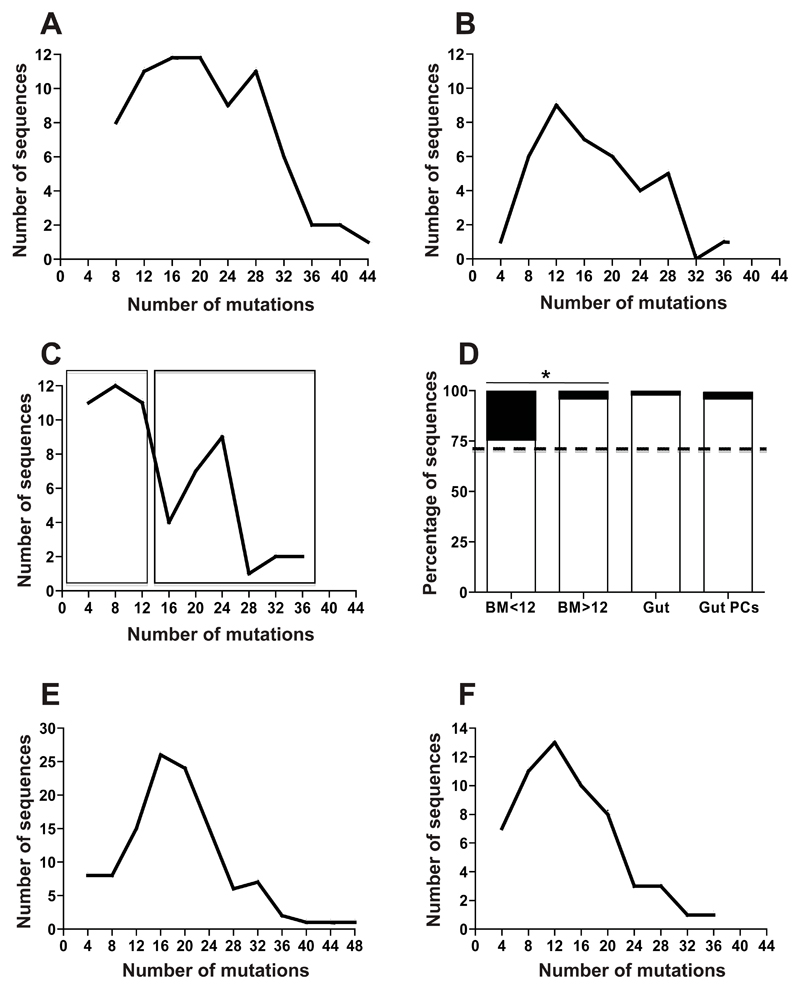
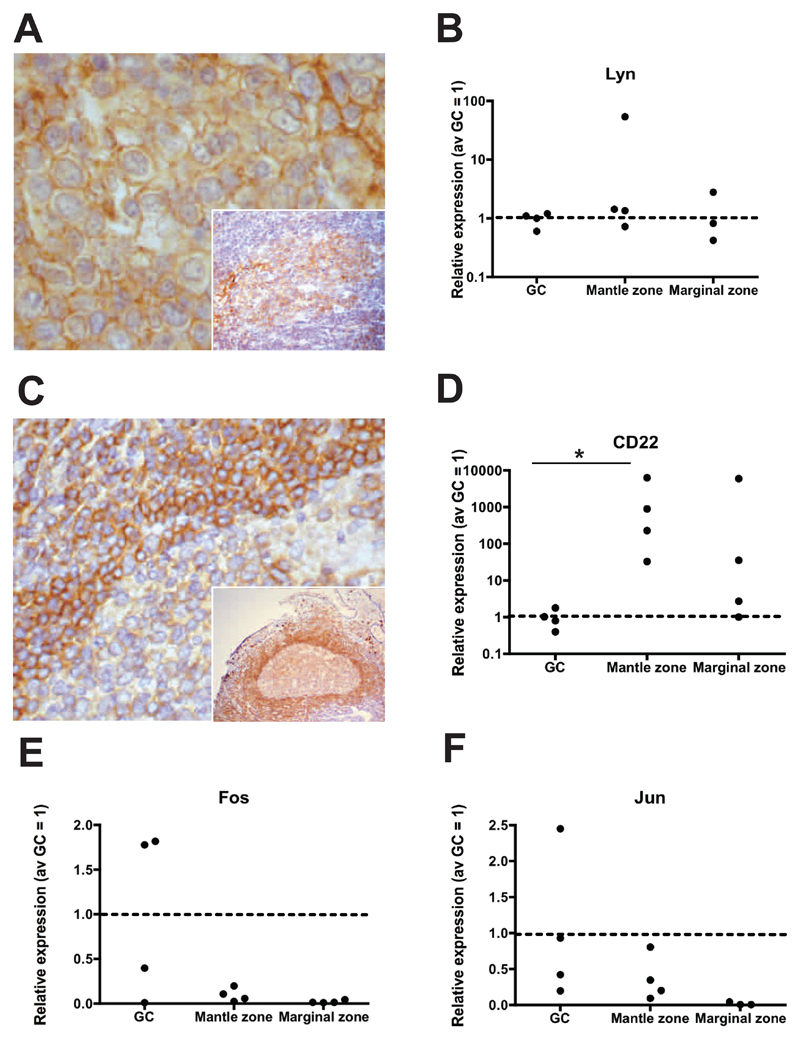
Results: Human intestinal IgA-producing plasma cells appeared to be of germinal center origin; there was no evidence for the population complexity that accompanies multiple pathways of derivation observed in bone marrow. In germinal center B cells of human GALT, Btk and Erk are phosphorylated, CD22 is down-regulated, Lyn is translocated to the cell membrane, and Fos and Jun are up-regulated; these features indicate B-cell receptor ligation during germinal center evolution. No differences in innate activation of B cells were observed in GALT, compared with peripheral immune compartments.
Conclusions: IgA-producing plasma cells appear to be derived from GALT germinal centers in humans. B-cell receptor engagement promotes formation of germinal centers of GALT, with no more evidence for innate immune receptor activation in the mucosa than nonintestinal immune compartments. Germinal centers in GALT should be targets of mucosal vaccinations because they are the source of human intestinal IgA response.
Copyright © 2011 AGA Institute. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare that there is no conflict of interest.
Figures
References
-
- Fagarasan S, Muramatsu M, Suzuki K, et al. Critical roles of activation-induced cytidine deaminase in the homeostasis of gut flora. Science. 2002;298:1424–7. - PubMed
-
- Braathen R, Sandvik A, Berntzen G, et al. Identification of a polymeric Ig receptor binding phage-displayed peptide that exploits epithelial transcytosis without dimeric IgA competition. J Biol Chem. 2006;281:7075–81. - PubMed
-
- Walker WA, Isselbacher KJ, Bloch KJ. Immunologic control of soluble protein absorption from the small intestine: a gut-surface phenomenon. Am J Clin Nutr. 1974;27:1434–40. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
