

Targeted JAM-C deletion in germ cells by Spo11-controlled Cre recombinase
- PMID: 21147852
- PMCID: PMC3001409
- DOI: 10.1242/jcs.072959
Targeted JAM-C deletion in germ cells by Spo11-controlled Cre recombinase
Abstract
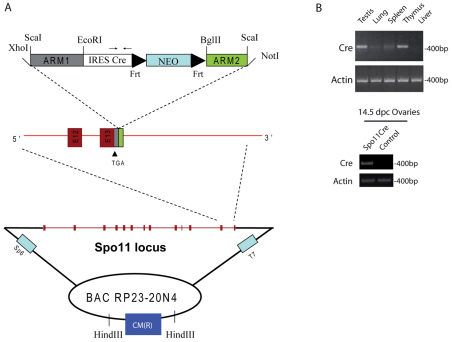
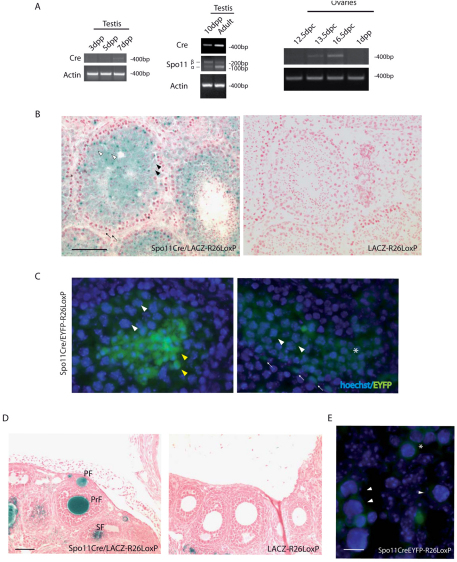
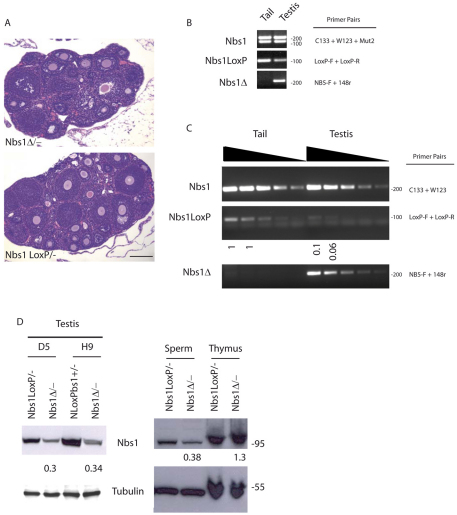
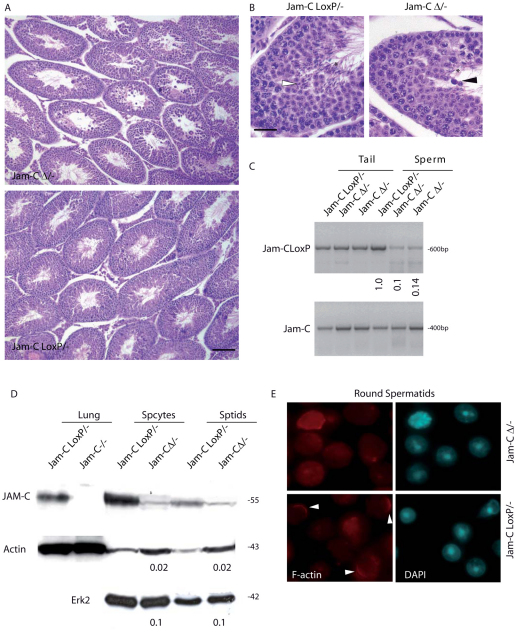
Meiosis is a crucial process for the production of functional gametes. However, the biological significance of many genes expressed during the meiotic phase remains poorly understood, mainly because of the lethal phenotypes of the knockout mice. Functional analysis of such genes using the conditional knockout approach is hindered by the lack of suitable Cre transgenic lines. We describe here the generation of transgenic mice expressing Cre recombinase under the control of the meiotic Spo11 gene. Using LacZ-R26(loxP) and EYFP-R26(loxP) reporter mice, we show the specific expression and activity of Cre during meiosis in males and females. Spo11(Cre) mice were then crossed with floxed Nbs1 and JAM-C mice to produce conditional knockouts. A strong reduction of Nbs1 and JAM-C protein levels was found in the testis. Although Nbs1-deleted mice developed minor gonadal abnormalities, JAM-C-knockout mice showed a spermiogenetic arrest, as previously described for the null mice. These results provide strong evidence that Spo11(Cre) transgenic mice represent a powerful tool for deleting genes of interest specifically in meiotic and/or in postmeiotic germ cells.
Figures
Similar articles
-
Postnatal male germ-cell expression of cre recombinase in Tex101-iCre transgenic mice.Genesis. 2010 Dec;48(12):717-22. doi: 10.1002/dvg.20675. Epub 2010 Oct 30. Genesis. 2010. PMID: 20853429 Free PMC article.
-
Transgenic mouse with high Cre recombinase activity in all prostate lobes, seminal vesicle, and ductus deferens.Prostate. 2003 Oct 1;57(2):160-4. doi: 10.1002/pros.10283. Prostate. 2003. PMID: 12949940
-
Incomplete cre-mediated excision leads to phenotypic differences between Stra8-iCre; Mov10l1(lox/lox) and Stra8-iCre; Mov10l1(lox/Δ) mice.Genesis. 2013 Jul;51(7):481-90. doi: 10.1002/dvg.22389. Epub 2013 Mar 30. Genesis. 2013. PMID: 23554062 Free PMC article.
-
Conditional gene targeting in the mouse nervous system: Insights into brain function and diseases.Pharmacol Ther. 2007 Mar;113(3):619-34. doi: 10.1016/j.pharmthera.2006.12.003. Epub 2007 Jan 10. Pharmacol Ther. 2007. PMID: 17289150 Review.
-
Mouse Cre Models for the Study of Bone Diseases.Curr Osteoporos Rep. 2018 Aug;16(4):466-477. doi: 10.1007/s11914-018-0455-7. Curr Osteoporos Rep. 2018. PMID: 29934753 Free PMC article. Review.
Cited by
-
Figla-Cre transgenic mice expressing myristoylated EGFP in germ cells provide a model for investigating perinatal oocyte dynamics.PLoS One. 2014 Jan 6;9(1):e84477. doi: 10.1371/journal.pone.0084477. eCollection 2014. PLoS One. 2014. PMID: 24400092 Free PMC article.
-
Adhesion Molecules Involved in Stem Cell Niche Retention During Normal Haematopoiesis and in Acute Myeloid Leukaemia.Front Immunol. 2021 Nov 12;12:756231. doi: 10.3389/fimmu.2021.756231. eCollection 2021. Front Immunol. 2021. PMID: 34867994 Free PMC article. Review.
-
A novel function of junctional adhesion molecule-C in mediating melanoma cell metastasis.Cancer Res. 2011 Jun 15;71(12):4096-105. doi: 10.1158/0008-5472.CAN-10-2794. Epub 2011 May 18. Cancer Res. 2011. PMID: 21593193 Free PMC article.
-
Junctional Adhesion Molecule 3 Expression in the Mouse Airway Epithelium Is Linked to Multiciliated Cells.Front Cell Dev Biol. 2021 Jul 28;9:622515. doi: 10.3389/fcell.2021.622515. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34395412 Free PMC article.
-
Genetic, structural, and chemical insights into the dual function of GRASP55 in germ cell Golgi remodeling and JAM-C polarized localization during spermatogenesis.PLoS Genet. 2017 Jun 15;13(6):e1006803. doi: 10.1371/journal.pgen.1006803. eCollection 2017 Jun. PLoS Genet. 2017. PMID: 28617811 Free PMC article.
References
-
- Ando H., Haruna Y., Miyazaki J., Okabe M., Nakanishi Y. (2000). Spermatocyte-specific gene excision by targeted expression of Cre recombinase. Biochem. Biophys. Res. Commun. 272, 125-128 - PubMed
-
- Barchi M., Geremia R., Magliozzi R., Bianchi E. (2009). Isolation and analyses of enriched populations of male mouse germ cells by sedimentation velocity: the centrifugal elutriation. Methods Mol. Biol. 558, 299-321 - PubMed
-
- Baudat F., Manova K., Yuen J. P., Jasin M., Keeney S. (2000). Chromosome synapsis defects and sexually dimorphic meiotic progression in mice lacking Spo11. Mol. Cell 6, 989-998 - PubMed
-
- Bellani M. A., Boateng K. A., Mc Leod D., Camerini-Otero R. D. (2010). The expression profile of the major mouse SPO11 isoforms indicates that SPO11β introduces the double strand breaks and suggests that SPO11α has an additional role in prophase both in spermatocytes and oocytes. Mol. Cell. Biol. 30, 4391-4403 - PMC - PubMed
-
- Bergqvist I., Eriksson B., Eriksson M., Holmberg D. (1998). Transgenic Cre recombinase expression in germ cells and early embryogenesis directs homogeneous and ubiquitous deletion of loxP-flanked gene segments. FEBS Lett. 438, 76-80 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
