

Cortical processing of dynamic sound envelope transitions
- PMID: 21148013
- PMCID: PMC3358036
- DOI: 10.1523/JNEUROSCI.2016-10.2010
Cortical processing of dynamic sound envelope transitions
Abstract
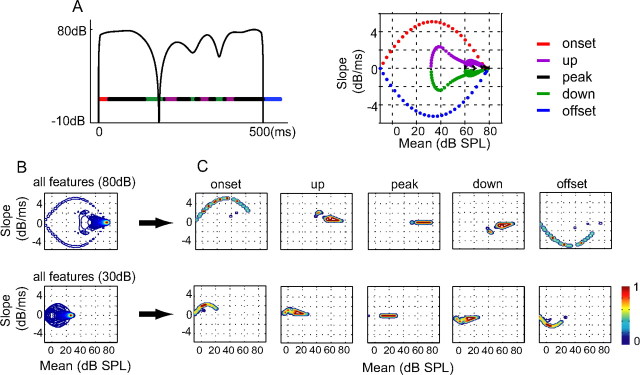
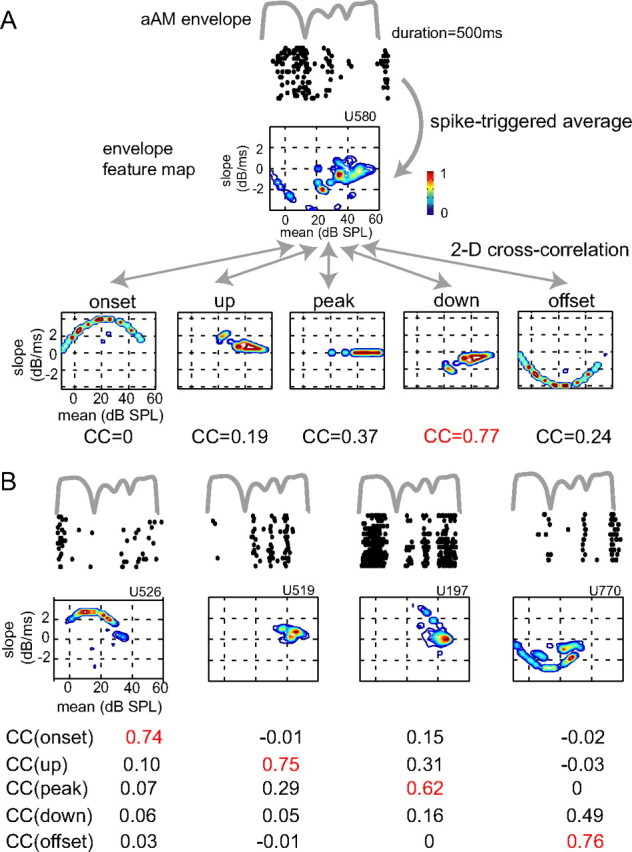
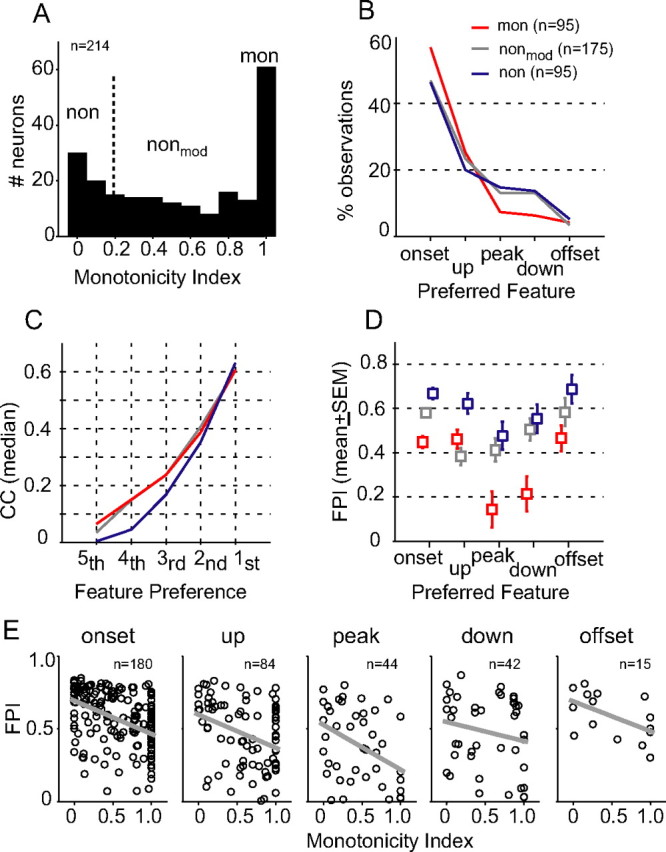
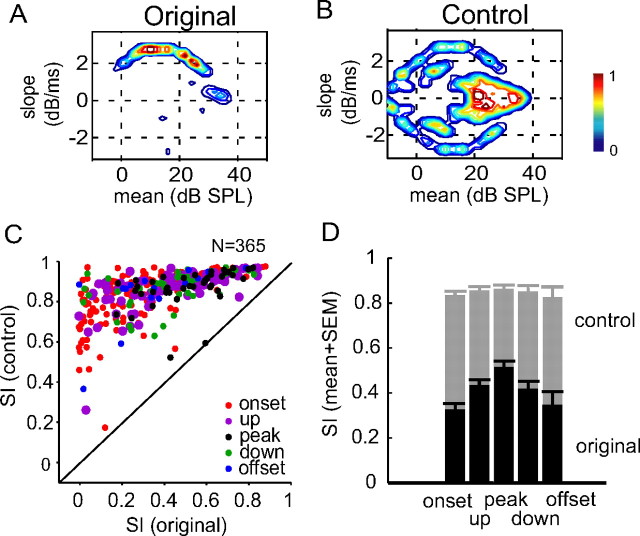
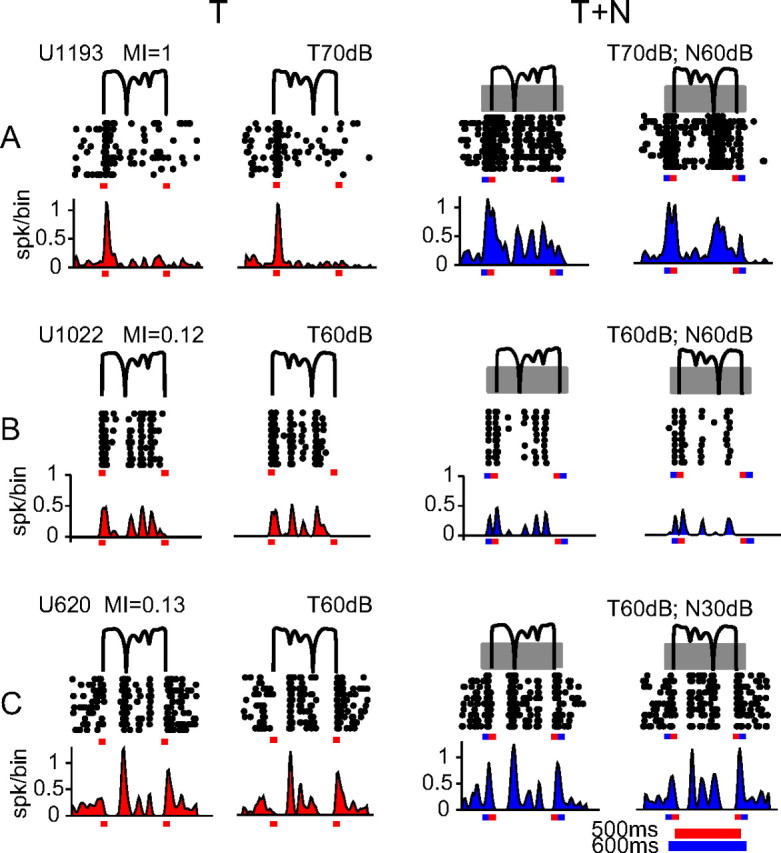
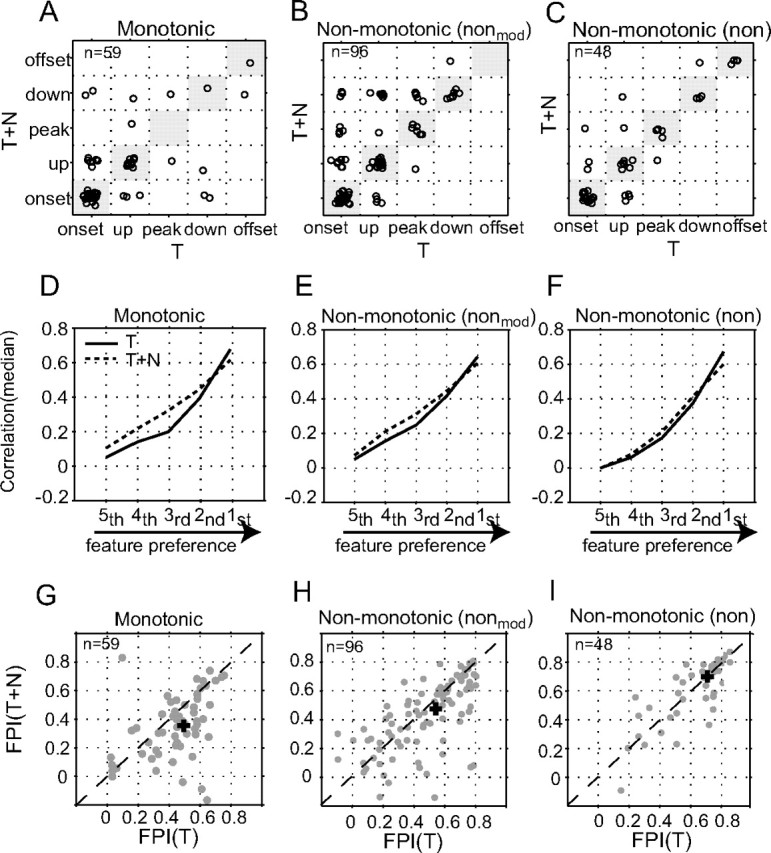
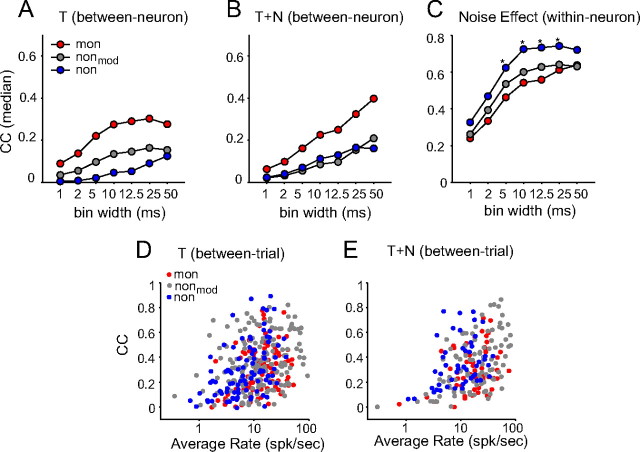
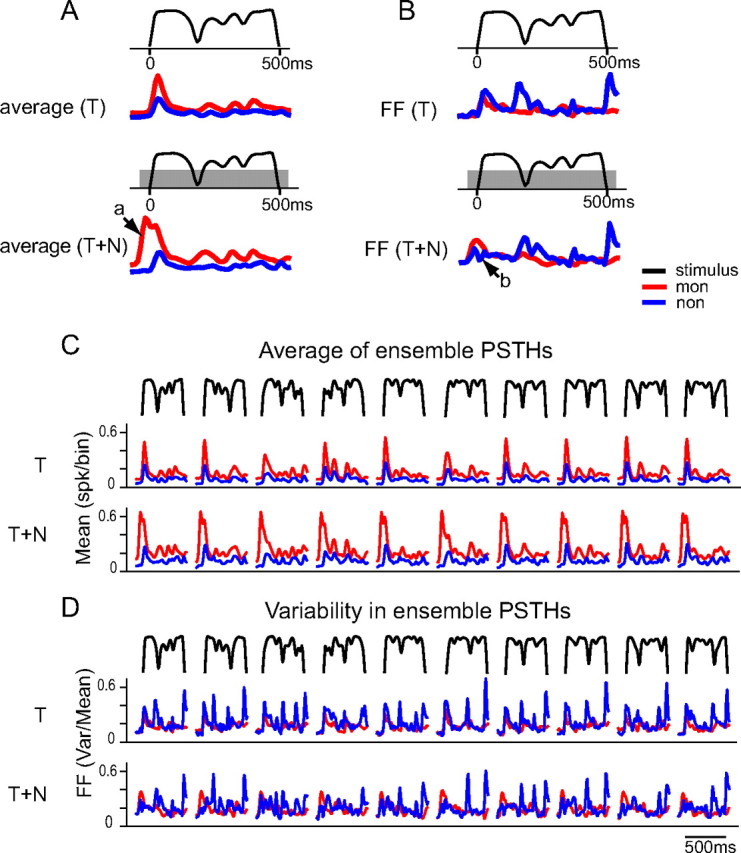
Slow envelope fluctuations in the range of 2-20 Hz provide important segmental cues for processing communication sounds. For a successful segmentation, a neural processor must capture envelope features associated with the rise and fall of signal energy, a process that is often challenged by the interference of background noise. This study investigated the neural representations of slowly varying envelopes in quiet and in background noise in the primary auditory cortex (A1) of awake marmoset monkeys. We characterized envelope features based on the local average and rate of change of sound level in envelope waveforms and identified envelope features to which neurons were selective by reverse correlation. Our results showed that envelope feature selectivity of A1 neurons was correlated with the degree of nonmonotonicity in their static rate-level functions. Nonmonotonic neurons exhibited greater feature selectivity than monotonic neurons in quiet and in background noise. The diverse envelope feature selectivity decreased spike-timing correlation among A1 neurons in response to the same envelope waveforms. As a result, the variability, but not the average, of the ensemble responses of A1 neurons represented more faithfully the dynamic transitions in low-frequency sound envelopes both in quiet and in background noise.
Figures
Similar articles
-
Encoding of sound envelope transients in the auditory cortex of juvenile rats and adult rats.Int J Dev Neurosci. 2016 Feb;48:50-7. doi: 10.1016/j.ijdevneu.2015.11.004. Epub 2015 Nov 26. Int J Dev Neurosci. 2016. PMID: 26626803
-
Temporal codes for amplitude contrast in auditory cortex.J Neurosci. 2010 Jan 13;30(2):767-84. doi: 10.1523/JNEUROSCI.4170-09.2010. J Neurosci. 2010. PMID: 20071542 Free PMC article.
-
Level invariant representation of sounds by populations of neurons in primary auditory cortex.J Neurosci. 2008 Mar 26;28(13):3415-26. doi: 10.1523/JNEUROSCI.2743-07.2008. J Neurosci. 2008. PMID: 18367608 Free PMC article.
-
How do auditory cortex neurons represent communication sounds?Hear Res. 2013 Nov;305:102-12. doi: 10.1016/j.heares.2013.03.011. Epub 2013 Apr 17. Hear Res. 2013. PMID: 23603138 Review.
-
Neural coding of temporal information in auditory thalamus and cortex.Neuroscience. 2008 Jun 12;154(1):294-303. doi: 10.1016/j.neuroscience.2008.03.065. Epub 2008 Apr 7. Neuroscience. 2008. Corrected and republished in: Neuroscience. 2008 Nov 19;157(2):484-94. doi: 10.1016/j.neuroscience.2008.07.050. PMID: 18555164 Free PMC article. Corrected and republished. Review.
Cited by
-
Neural correlates of auditory scene analysis and perception.Int J Psychophysiol. 2015 Feb;95(2):238-245. doi: 10.1016/j.ijpsycho.2014.03.004. Epub 2014 Mar 25. Int J Psychophysiol. 2015. PMID: 24681354 Free PMC article. Review.
-
Hearing the light: neural and perceptual encoding of optogenetic stimulation in the central auditory pathway.Sci Rep. 2015 May 22;5:10319. doi: 10.1038/srep10319. Sci Rep. 2015. PMID: 26000557 Free PMC article.
-
Thresholding of auditory cortical representation by background noise.Front Neural Circuits. 2014 Nov 10;8:133. doi: 10.3389/fncir.2014.00133. eCollection 2014. Front Neural Circuits. 2014. PMID: 25426029 Free PMC article.
-
Decision making as a window on cognition.Neuron. 2013 Oct 30;80(3):791-806. doi: 10.1016/j.neuron.2013.10.047. Neuron. 2013. PMID: 24183028 Free PMC article. Review.
-
Neural populations within macaque early vestibular pathways are adapted to encode natural self-motion.PLoS Biol. 2024 Apr 30;22(4):e3002623. doi: 10.1371/journal.pbio.3002623. eCollection 2024 Apr. PLoS Biol. 2024. PMID: 38687807 Free PMC article.
References
-
- Bieser A, Müller-Preuss P. Auditory responsive cortex in the squirrel monkey: neural responses to amplitude-modulated sounds. Exp Brain Res. 1996;108:273–284. - PubMed
-
- Bregman AS. Auditory scene analysis: the perceptual organization of sound. Cambridge: MIT; 1990.
-
- Cariani P. As if time really mattered: temporal strategies for neural coding of sensory information. Commun Cogn Artif Intell. 1995;12:161–229.
-
- de Boer E, de Jongh HR. On cochlear encoding: potentialities and limitations of the reverse-correlation technique. J Acoust Soc Am. 1978;63:115–135. - PubMed
-
- deCharms RC, Merzenich MM. Primary cortical representation of sounds by the coordination of action-potential timing. Nature. 1996;381:610–613. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials