

Recombination can cause telomere elongations as well as truncations deep within telomeres in wild-type Kluyveromyces lactis cells
- PMID: 21148753
- PMCID: PMC3067406
- DOI: 10.1128/EC.00209-10
Recombination can cause telomere elongations as well as truncations deep within telomeres in wild-type Kluyveromyces lactis cells
Abstract
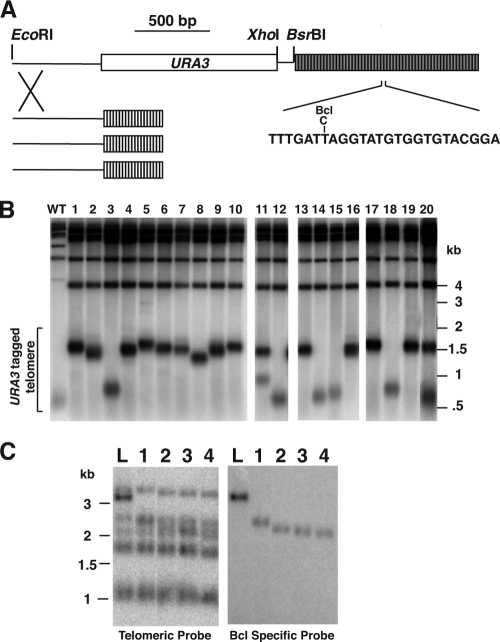
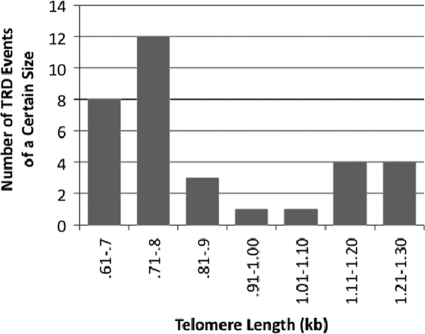
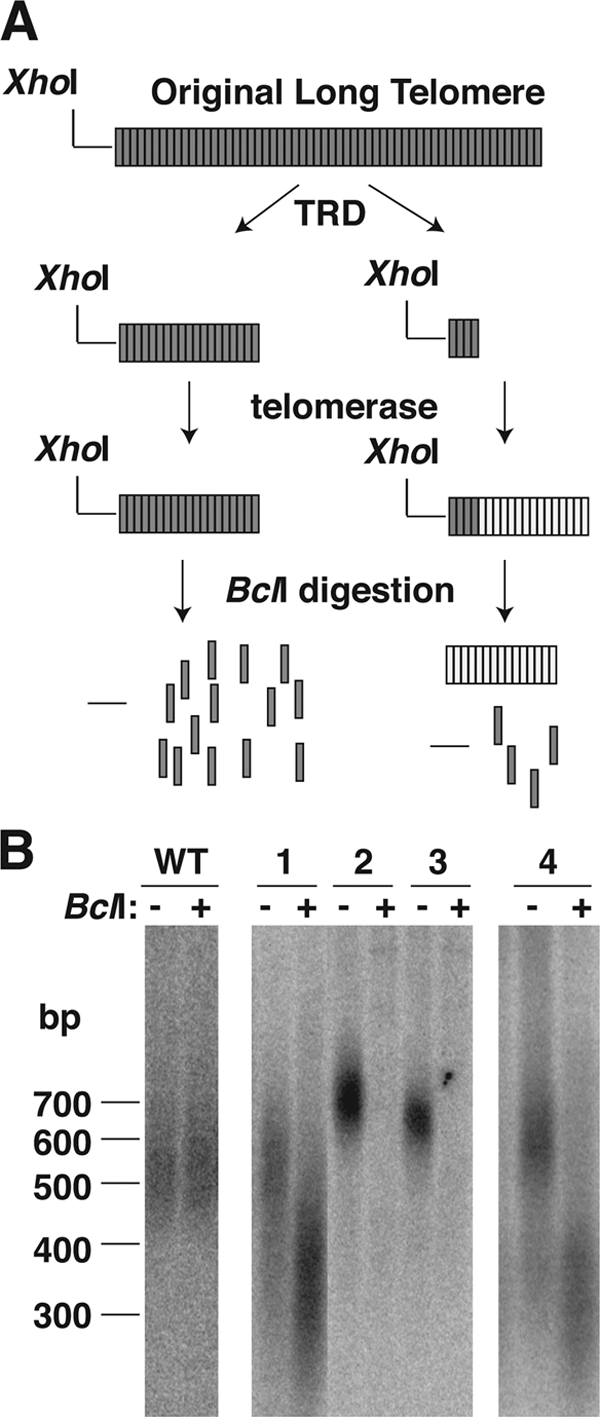
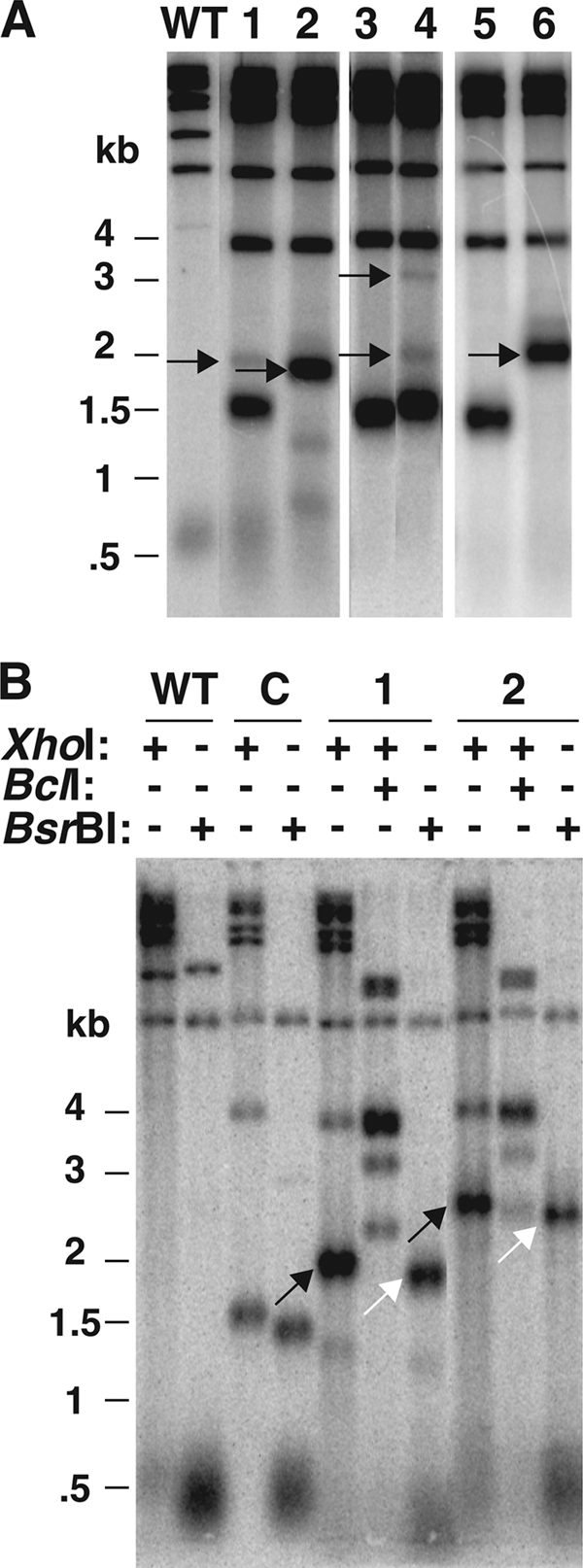
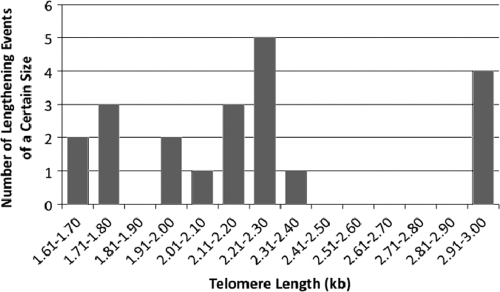
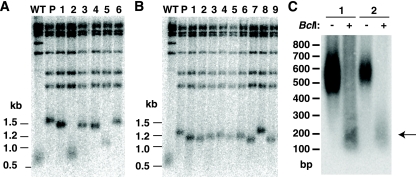
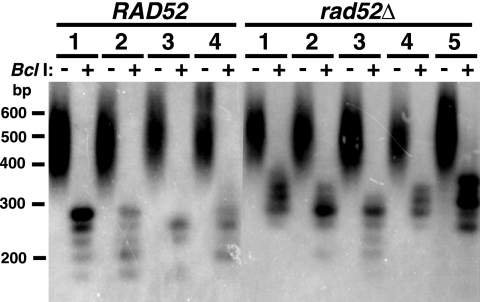
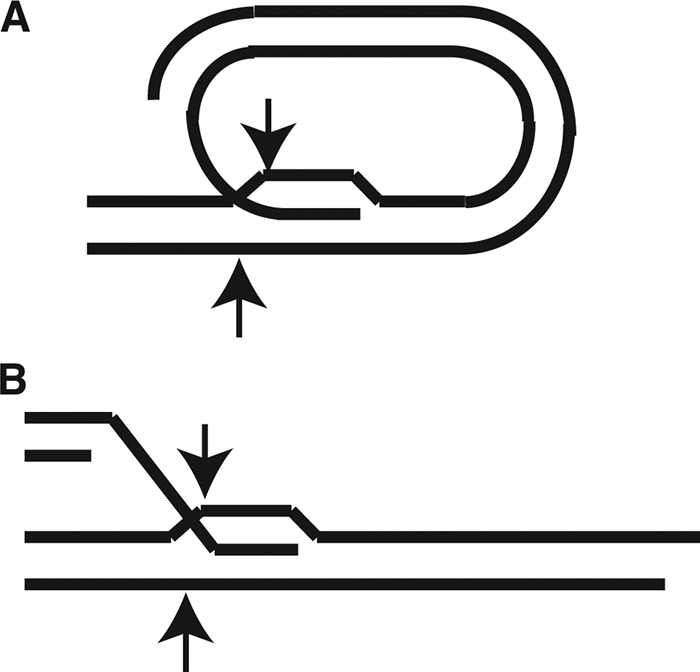
In this study, we examined the role of recombination at the telomeres of the yeast Kluyveromyces lactis. We demonstrated that an abnormally long and mutationally tagged telomere was subject to high rates of telomere rapid deletion (TRD) that preferentially truncated the telomere to near-wild-type size. Unlike the case in Saccharomyces cerevisiae, however, there was not a great increase in TRD in meiosis. About half of mitotic TRD events were associated with deep turnover of telomeric repeats, suggesting that telomeres were often cleaved to well below normal length prior to being reextended by telomerase. Despite its high rate of TRD, the long telomere showed no increase in the rate of subtelomeric gene conversion, a highly sensitive test of telomere dysfunction. We also showed that the long telomere was subject to appreciable rates of becoming elongated substantially further through a recombinational mechanism that added additional tagged repeats. Finally, we showed that the deep turnover that occurs within normal-length telomeres was diminished in the absence of RAD52. Taken together, our results suggest that homologous recombination is a significant process acting on both abnormally long and normally sized telomeres in K. lactis.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
