

Mechanistic insights into an engineered riboswitch: a switching element which confers riboswitch activity
- PMID: 21149263
- PMCID: PMC3082870
- DOI: 10.1093/nar/gkq946
Mechanistic insights into an engineered riboswitch: a switching element which confers riboswitch activity
Abstract
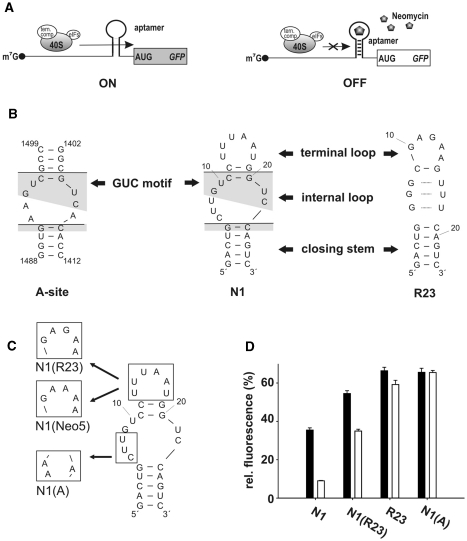
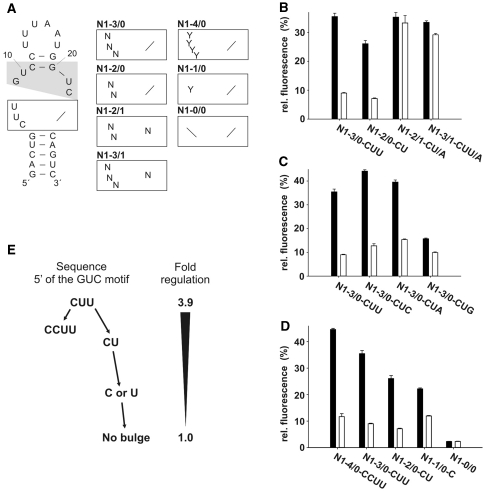
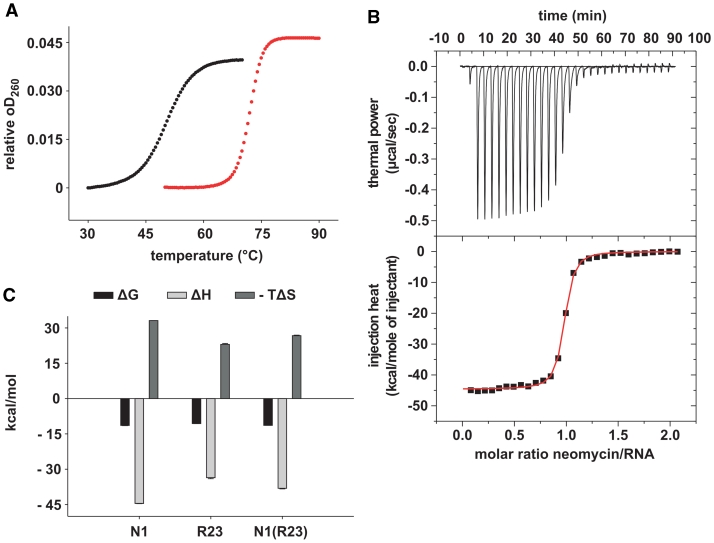
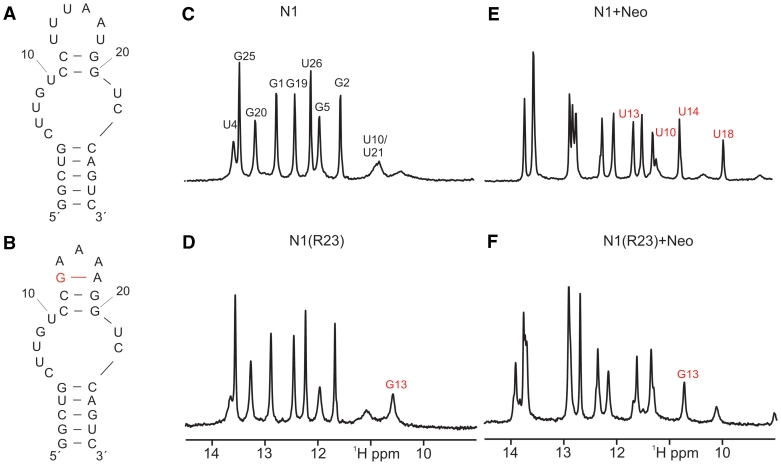
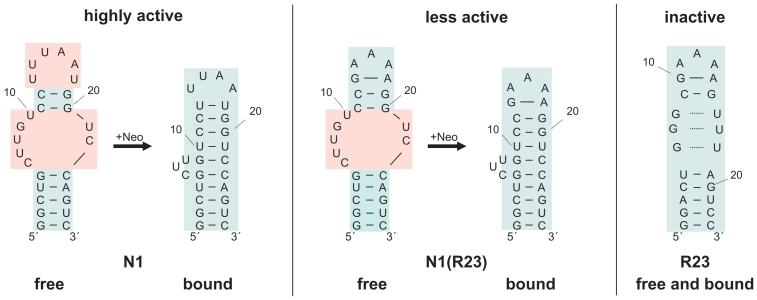
While many different RNA aptamers have been identified that bind to a plethora of small molecules only very few are capable of acting as engineered riboswitches. Even for aptamers binding the same ligand large differences in their regulatory potential were observed. We address here the molecular basis for these differences by using a set of unrelated neomycin-binding aptamers. UV melting analyses showed that regulating aptamers are thermally stabilized to a significantly higher degree upon ligand binding than inactive ones. Regulating aptamers show high ligand-binding affinity in the low nanomolar range which is necessary but not sufficient for regulation. NMR data showed that a destabilized, open ground state accompanied by extensive structural changes upon ligand binding is important for regulation. In contrast, inactive aptamers are already pre-formed in the absence of the ligand. By a combination of genetic, biochemical and structural analyses, we identified a switching element responsible for destabilizing the ligand free state without compromising the bound form. Our results explain for the first time the molecular mechanism of an engineered riboswitch.
Figures
References
-
- Suess B, Weigand JE. Engineered riboswitches: overview, problems and trends. RNA Biol. 2008;5:24–29. - PubMed
-
- Weigand JE, Suess B. Aptamers and riboswitches: perspectives in biotechnology. Appl. Microbiol. Biotechnol. 2009;85:229–236. - PubMed
-
- Hermann T, Patel DJ. Adaptive recognition by nucleic acid aptamers. Science. 2000;287:820–825. - PubMed
-
- Hanson S, Berthelot K, Fink B, McCarthy JE, Suess B. Tetracycline-aptamer-mediated translational regulation in yeast. Mol. Microbiol. 2003;49:1627–1637. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
