

T-cell cytokines differentially control human monocyte antimicrobial responses by regulating vitamin D metabolism
- PMID: 21149724
- PMCID: PMC3012480
- DOI: 10.1073/pnas.1011624108
T-cell cytokines differentially control human monocyte antimicrobial responses by regulating vitamin D metabolism
Abstract
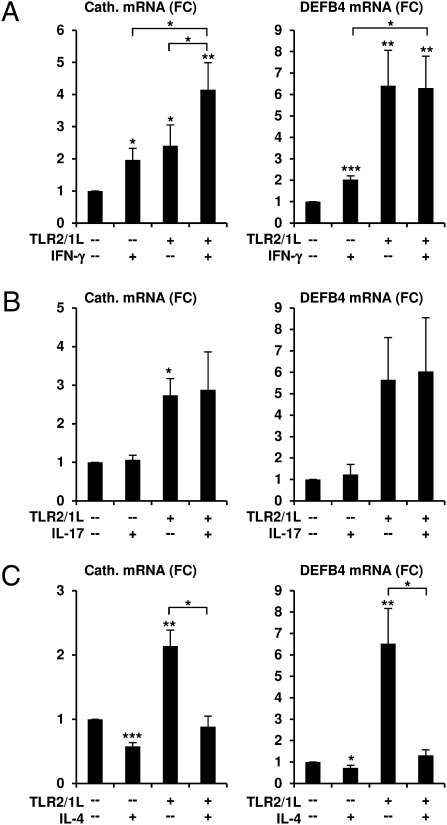
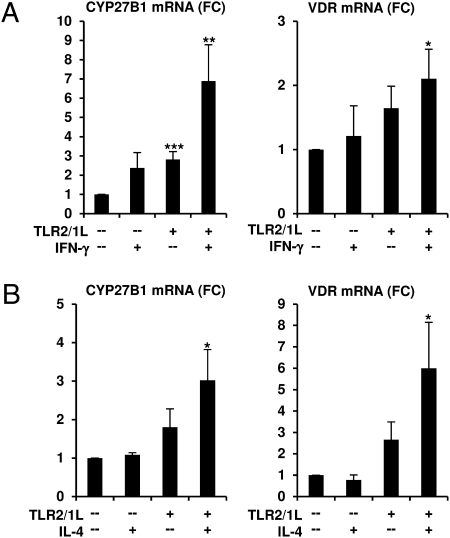
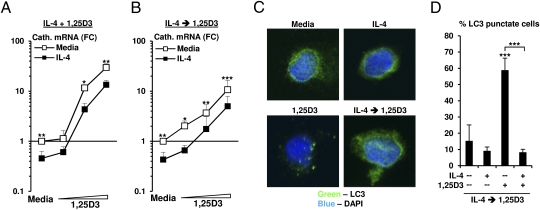
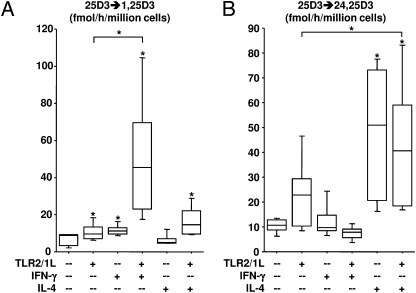
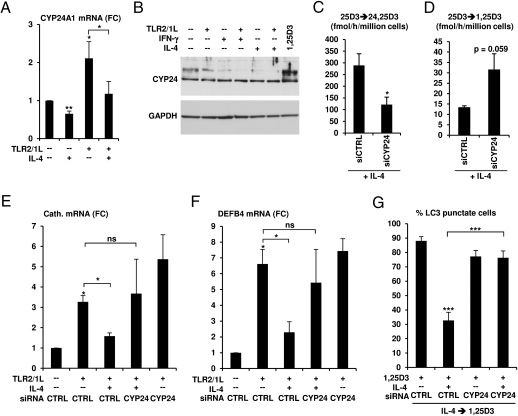
We investigated the mechanisms by which T-cell cytokines are able to influence the Toll-like receptor (TLR)-induced, vitamin D-dependent antimicrobial pathway in human monocytes. T-cell cytokines differentially influenced TLR2/1-induced expression of the antimicrobial peptides cathelicidin and DEFB4, being up-regulated by IFN-γ, down-regulated by IL-4, and unaffected by IL-17. The Th1 cytokine IFN-γ up-regulated TLR2/1 induction of 25-hydroxyvitamin D-1α-hydroxylase (i.e., CYP27B1), leading to enhanced bioconversion of 25-hydroxyvitamin D(3) (25D(3)) to its active metabolite 1,25D(3). In contrast, the Th2 cytokine IL-4, by itself and in combination with the TLR2/1 ligand, induced catabolism of 25D(3) to the inactive metabolite 24,25D(3), and was dependent on expression of vitamin D-24-hydroxylase (i.e., CYP24A1). Therefore, the ability of T-cell cytokines to differentially control monocyte vitamin D metabolism represents a mechanism by which cell-mediated immune responses can regulate innate immune mechanisms to defend against microbial pathogens.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Brightbill HD, et al. Host defense mechanisms triggered by microbial lipoproteins through toll-like receptors. Science. 1999;285:732–736. - PubMed
-
- Aliprantis AO, et al. Cell activation and apoptosis by bacterial lipoproteins through toll-like receptor-2. Science. 1999;285:736–739. - PubMed
-
- Takeuchi O, et al. Role of TLR1 in mediating immune response to microbial lipoproteins. J Immunol. 2002;169:10–14. - PubMed
-
- Thoma-Uszynski S, et al. Induction of direct antimicrobial activity through mammalian toll-like receptors. Science. 2001;291:1544–1547. - PubMed
-
- Wang TT, et al. Cutting edge: 1,25-dihydroxyvitamin D3 is a direct inducer of antimicrobial peptide gene expression. J Immunol. 2004;173:2909–2912. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
