

NER and HR pathways act sequentially to promote UV-C-induced germ cell apoptosis in Caenorhabditis elegans
- PMID: 21151025
- PMCID: PMC3131928
- DOI: 10.1038/cdd.2010.158
NER and HR pathways act sequentially to promote UV-C-induced germ cell apoptosis in Caenorhabditis elegans
Abstract
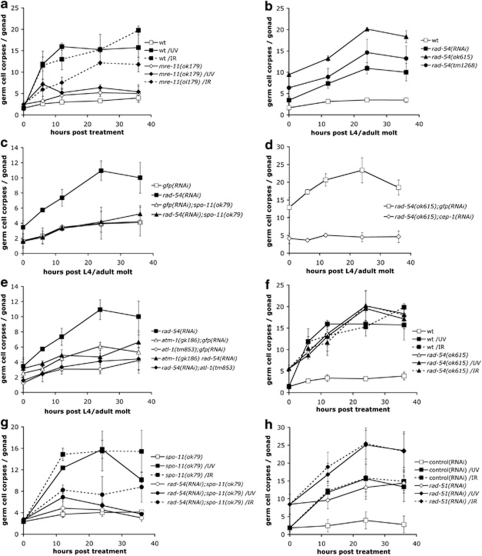
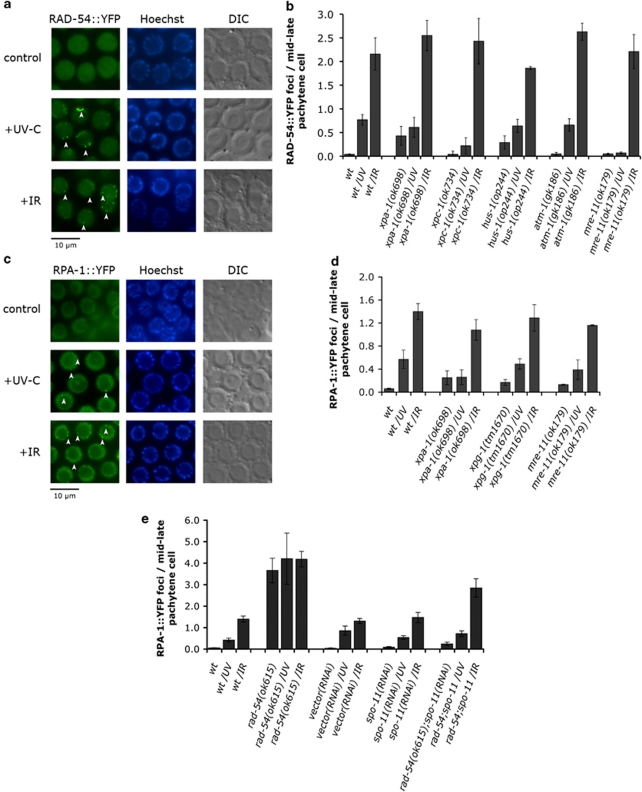
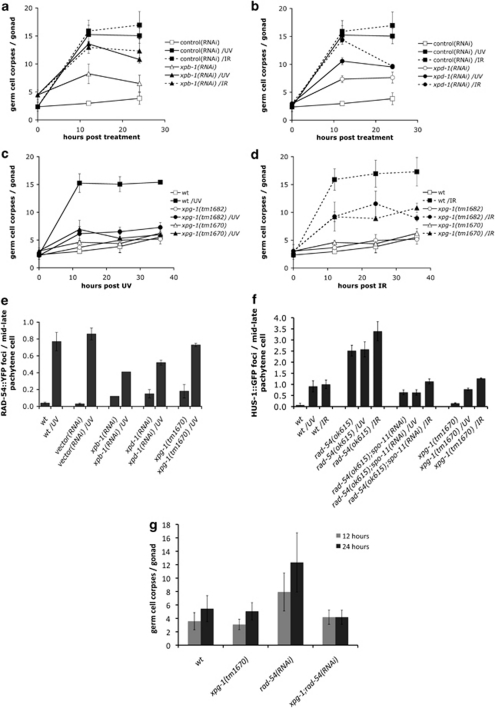
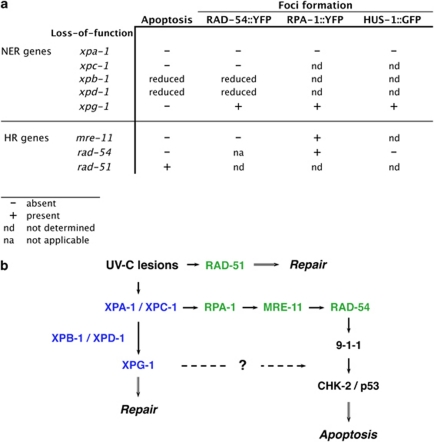
Ultraviolet (UV) radiation-induced DNA damage evokes a complex network of molecular responses, which culminate in DNA repair, cell cycle arrest and apoptosis. Here, we provide an in-depth characterization of the molecular pathway that mediates UV-C-induced apoptosis of meiotic germ cells in the nematode Caenorhabditis elegans. We show that UV-C-induced DNA lesions are not directly pro-apoptotic. Rather, they must first be recognized and processed by the nucleotide excision repair (NER) pathway. Our data suggest that NER pathway activity transforms some of these lesions into other types of DNA damage, which in turn are recognized and acted upon by the homologous recombination (HR) pathway. HR pathway activity is in turn required for the recruitment of the C. elegans homolog of the yeast Rad9-Hus1-Rad1 (9-1-1) complex and activation of downstream checkpoint kinases. Blocking either the NER or HR pathway abrogates checkpoint pathway activation and UV-C-induced apoptosis. Our results show that, following UV-C, multiple DNA repair pathways can cooperate to signal to the apoptotic machinery to eliminate potentially hazardous cells.
Figures

Similar articles
-
The nucleotide excision repair pathway is required for UV-C-induced apoptosis in Caenorhabditis elegans.Cell Death Differ. 2007 Jun;14(6):1129-38. doi: 10.1038/sj.cdd.4402115. Epub 2007 Mar 9. Cell Death Differ. 2007. PMID: 17347667
-
Methods for studying the DNA damage response in the Caenorhabdatis elegans germ line.Methods Cell Biol. 2012;107:321-52. doi: 10.1016/B978-0-12-394620-1.00011-4. Methods Cell Biol. 2012. PMID: 22226529 Review.
-
A conserved checkpoint pathway mediates DNA damage--induced apoptosis and cell cycle arrest in C. elegans.Mol Cell. 2000 Mar;5(3):435-43. doi: 10.1016/s1097-2765(00)80438-4. Mol Cell. 2000. PMID: 10882129
-
ZTF-8 interacts with the 9-1-1 complex and is required for DNA damage response and double-strand break repair in the C. elegans germline.PLoS Genet. 2014 Oct 16;10(10):e1004723. doi: 10.1371/journal.pgen.1004723. eCollection 2014 Oct. PLoS Genet. 2014. PMID: 25329393 Free PMC article.
-
Death and more: DNA damage response pathways in the nematode C. elegans.Cell Death Differ. 2004 Jan;11(1):21-8. doi: 10.1038/sj.cdd.4401340. Cell Death Differ. 2004. PMID: 14685168 Review.
Cited by
-
Combination of Aβ Secretion and Oxidative Stress in an Alzheimer-Like Cell Line Leads to the Over-Expression of the Nucleotide Excision Repair Proteins DDB2 and XPC.Int J Mol Sci. 2015 Jul 30;16(8):17422-44. doi: 10.3390/ijms160817422. Int J Mol Sci. 2015. PMID: 26263968 Free PMC article.
-
Differential regulation of DNA damage response activation between somatic and germline cells in Caenorhabditis elegans.Cell Death Differ. 2012 Nov;19(11):1847-55. doi: 10.1038/cdd.2012.69. Epub 2012 Jun 15. Cell Death Differ. 2012. PMID: 22705849 Free PMC article.
-
The unfolded protein response of the endoplasmic reticulum protects Caenorhabditis elegans against DNA damage caused by stalled replication forks.G3 (Bethesda). 2024 Apr 3;14(4):jkae017. doi: 10.1093/g3journal/jkae017. G3 (Bethesda). 2024. PMID: 38267027 Free PMC article.
-
Ovarian cancer resistance to PARPi and platinum-containing chemotherapy.Cancer Drug Resist. 2022 Jun 22;5(3):637-646. doi: 10.20517/cdr.2021.146. eCollection 2022. Cancer Drug Resist. 2022. PMID: 36176755 Free PMC article.
-
Caenorhabditis elegans as a Model to Study Aging and Photoaging.Biomolecules. 2024 Sep 30;14(10):1235. doi: 10.3390/biom14101235. Biomolecules. 2024. PMID: 39456168 Free PMC article. Review.
References
-
- Hanahan D, Weinberg RA. The hallmarks of cancer. Cell. 2000;100:57–70. - PubMed
-
- Bartek J, Lukas J. DNA damage checkpoints: from initiation to recovery or adaptation. Curr Opin Cell Biol. 2007;19:238–245. - PubMed
-
- Sancar A, Lindsey-Boltz LA, Unsal-Kacmaz K, Linn S. Molecular mechanisms of mammalian DNA repair and the DNA damage checkpoints. Annu Rev Biochem. 2004;73:39–85. - PubMed
-
- Cleaver JE. Cancer in xeroderma pigmentosum and related disorders of DNA repair. Nat Rev Cancer. 2005;5:564–573. - PubMed
-
- de Boer J, Hoeijmakers JH. Cancer from the outside, aging from the inside: mouse models to study the consequences of defective nucleotide excision repair. Biochimie. 1999;81:127–137. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
