

Directed differentiation of human pluripotent stem cells into intestinal tissue in vitro
- PMID: 21151107
- PMCID: PMC3033971
- DOI: 10.1038/nature09691
Directed differentiation of human pluripotent stem cells into intestinal tissue in vitro
Abstract
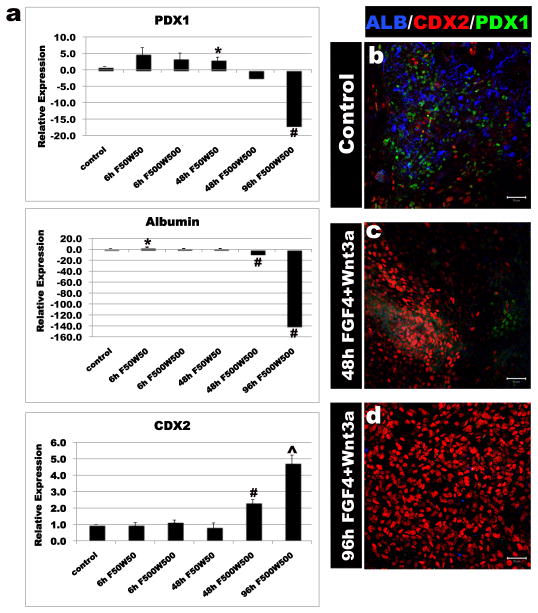
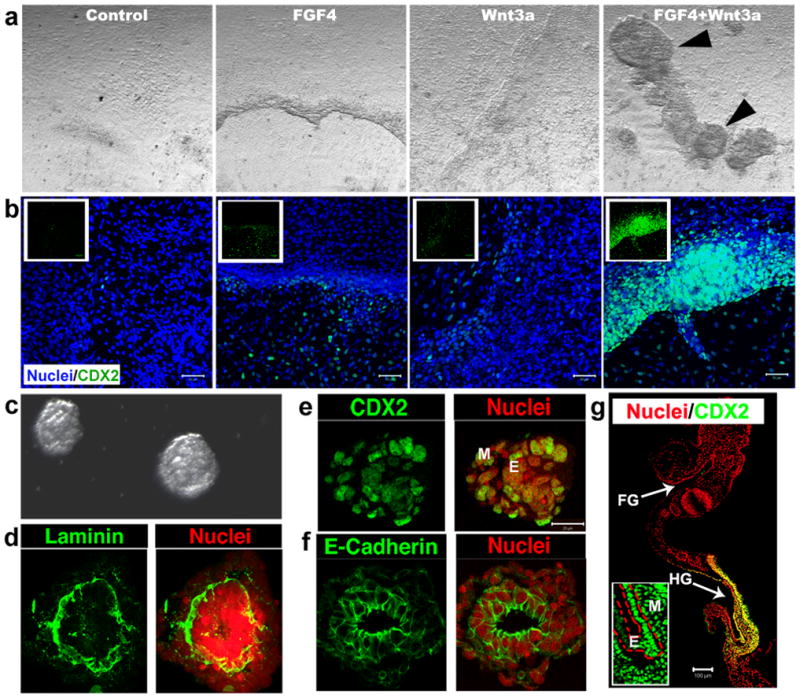
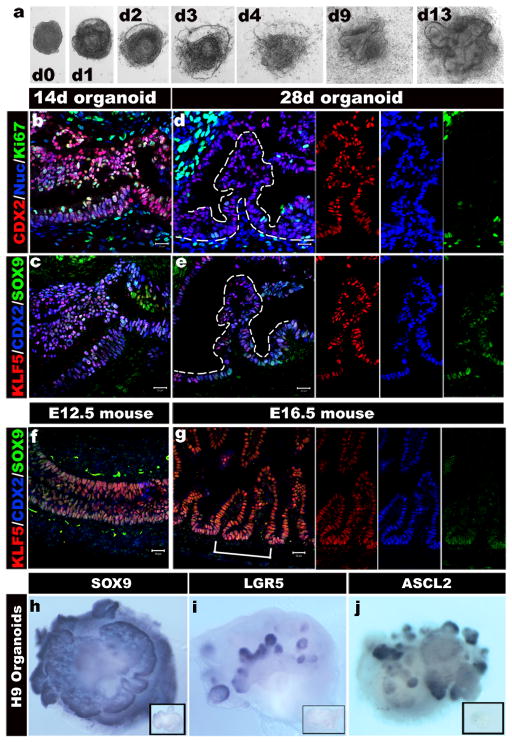
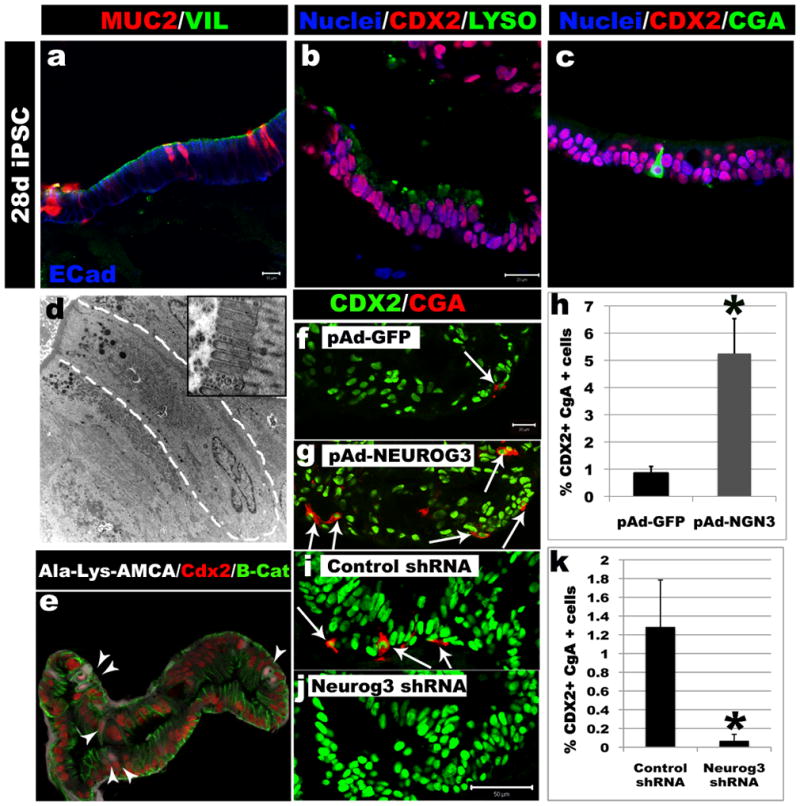
Studies in embryonic development have guided successful efforts to direct the differentiation of human embryonic and induced pluripotent stem cells (PSCs) into specific organ cell types in vitro. For example, human PSCs have been differentiated into monolayer cultures of liver hepatocytes and pancreatic endocrine cells that have therapeutic efficacy in animal models of liver disease and diabetes, respectively. However, the generation of complex three-dimensional organ tissues in vitro remains a major challenge for translational studies. Here we establish a robust and efficient process to direct the differentiation of human PSCs into intestinal tissue in vitro using a temporal series of growth factor manipulations to mimic embryonic intestinal development. This involved activin-induced definitive endoderm formation, FGF/Wnt-induced posterior endoderm pattering, hindgut specification and morphogenesis, and a pro-intestinal culture system to promote intestinal growth, morphogenesis and cytodifferentiation. The resulting three-dimensional intestinal 'organoids' consisted of a polarized, columnar epithelium that was patterned into villus-like structures and crypt-like proliferative zones that expressed intestinal stem cell markers. The epithelium contained functional enterocytes, as well as goblet, Paneth and enteroendocrine cells. Using this culture system as a model to study human intestinal development, we identified that the combined activity of WNT3A and FGF4 is required for hindgut specification whereas FGF4 alone is sufficient to promote hindgut morphogenesis. Our data indicate that human intestinal stem cells form de novo during development. We also determined that NEUROG3, a pro-endocrine transcription factor that is mutated in enteric anendocrinosis, is both necessary and sufficient for human enteroendocrine cell development in vitro. PSC-derived human intestinal tissue should allow for unprecedented studies of human intestinal development and disease.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
Comment in
-
Differentiation in three dimensions.Nat Methods. 2011 Feb;8(2):111. doi: 10.1038/nmeth0211-111. Nat Methods. 2011. PMID: 21355124 No abstract available.
-
Stem cells: Intestinal tissue generated in vitro.Nat Rev Gastroenterol Hepatol. 2011 Feb;8(2):63. doi: 10.1038/nrgastro.2010.226. Nat Rev Gastroenterol Hepatol. 2011. PMID: 21381243 No abstract available.
References
-
- Spence JR, Wells JM. Translational embryology: Using embryonic principles to generate pancreatic endocrine cells from embryonic stem cells. Developmental Dynamics. 2007;236:3218–3227. - PubMed
-
- Cai J, et al. Directed differentiation of human embryonic stem cells into functional hepatic cells. Hepatology. 2007;45:1229–1239. - PubMed
-
- D’Amour KA, et al. Production of pancreatic hormone-expressing endocrine cells from human embryonic stem cells. Nat Biotechnol. 2006;24:1392–1401. - PubMed
-
- Song Z, et al. Efficient generation of hepatocyte-like cells from human induced pluripotent stem cells. Cell research. 2009;19:1233–1242. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- F32 DK83202-01/DK/NIDDK NIH HHS/United States
- T32 HD007463/HD/NICHD NIH HHS/United States
- R01 CA142826/CA/NCI NIH HHS/United States
- P30 DK078392/DK/NIDDK NIH HHS/United States
- T32 HD07463/HD/NICHD NIH HHS/United States
- R01 DK080823/DK/NIDDK NIH HHS/United States
- UL1 TR000077/TR/NCATS NIH HHS/United States
- U54 RR025216/RR/NCRR NIH HHS/United States
- R03 DK084167/DK/NIDDK NIH HHS/United States
- R01GM072915/GM/NIGMS NIH HHS/United States
- R01DK080823A1/DK/NIDDK NIH HHS/United States
- R01 GM072915/GM/NIGMS NIH HHS/United States
- R01 DK092456/DK/NIDDK NIH HHS/United States
- F32 DK083202/DK/NIDDK NIH HHS/United States
- K01 DK091415/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
