

What do we gain from gamma? Local dynamic gain modulation drives enhanced efficacy and efficiency of signal transmission
- PMID: 21151350
- PMCID: PMC2981421
- DOI: 10.3389/fnhum.2010.00185
What do we gain from gamma? Local dynamic gain modulation drives enhanced efficacy and efficiency of signal transmission
Abstract
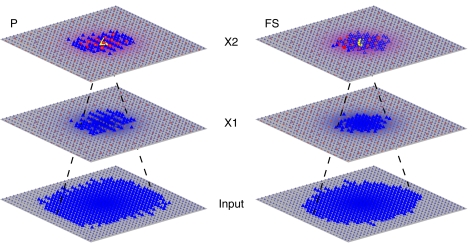
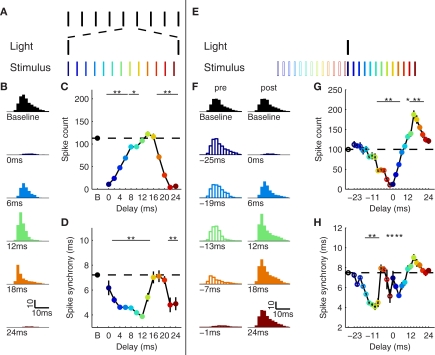
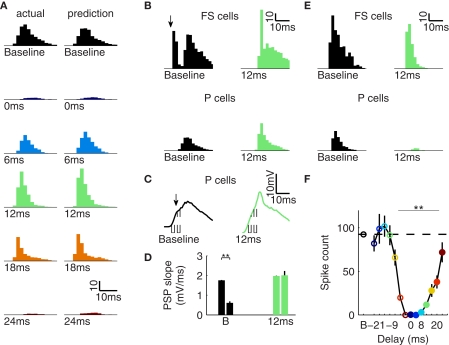
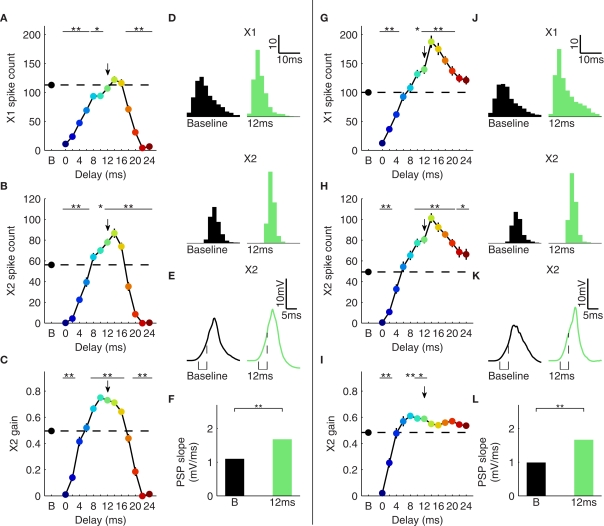
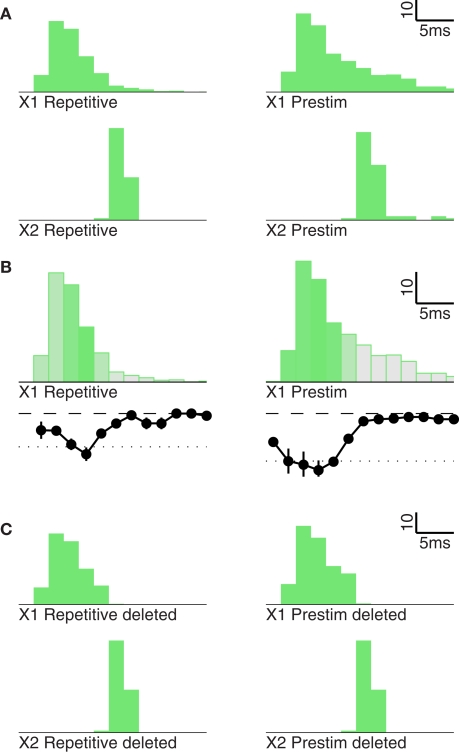
Gamma oscillations in neocortex are hypothesized to improve information transmission between groups of neurons. We recently showed that optogenetic drive of fast-spiking interneurons (FS) at 40 Hz in mouse neocortex in vivo modulates the spike count and precision of sensory evoked responses. At specific phases of alignment between stimuli and FS activation, total evoked spike count was unchanged compared to baseline, but precision was increased. In the present study, we used computational modeling to investigate the origin of these local transformations, and to make predictions about their impact on downstream signal transmission. We replicated the prior experimental findings, and found that the local gain observed can be explained by mutual inhibition of fast-spiking interneurons, leading to more robust sensory-driven spiking in a brief temporal window post-stimulus, increasing local synchrony. Enhanced spiking in a second neocortical area, without a net increase in overall driven spikes in the first area, resulted from faster depolarization of target neurons due to increased pre-synaptic synchrony. In addition, we found that the precise temporal structure of spiking in the first area impacted the gain between cortical areas. The optimal spike distribution matched the "window of opportunity" defined by the timing of inhibition in the target area: spiking beyond this window did not contribute to downstream spike generation, leading to decreased overall gain. This result predicts that efficient transmission between neocortical areas requires a mechanism to dynamically match the temporal structure of the output of one area to the timing of inhibition in the recipient zone.
Keywords: cortex; gain; gamma; inhibition; model; sensory; synchrony.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources