

Long-Range GABAergic Connections Distributed throughout the Neocortex and their Possible Function
- PMID: 21151790
- PMCID: PMC3000116
- DOI: 10.3389/fnins.2010.00202
Long-Range GABAergic Connections Distributed throughout the Neocortex and their Possible Function
Abstract
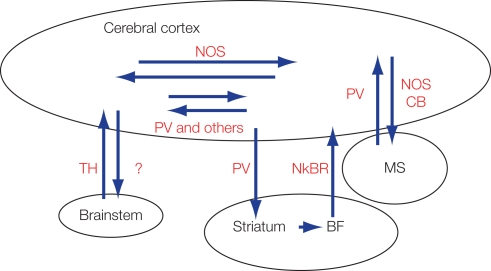
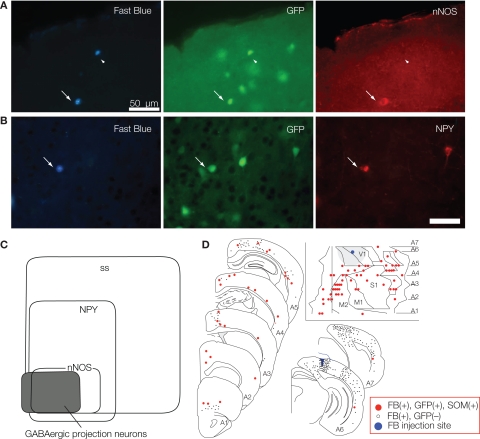
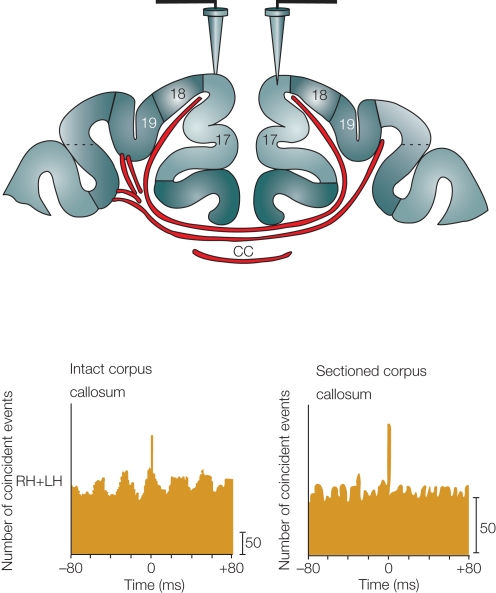
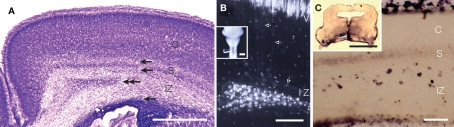
Features and functions of long-range GABAergic projection neurons in the developing cerebral cortex have been reported previously, although until now their significance in the adult cerebral cortex has remained uncertain. The septo-hippocampal circuit is one exception - in this system, long-range mature GABAergic projection neurons have been well analyzed and their contribution to the generation of theta-oscillatory behavior in the hippocampus has been documented. To have a clue to the function of the GABAergic projection neurons in the neocortex, we view how the long-range GABAergic projections are integrated in the cortico-cortical, cortico-fugal, and afferent projections in the cerebral cortex. Then, we consider the possibility that the GABAergic projection neurons are involved in the generation, modification, and/or synchronization of oscillations in mature neocortical neuron activity. When markers that identify the GABAergic projection neurons are examined in anatomical and developmental studies, it is clear that neuronal NO synthetase (nNOS)-immunoreactivity can readily identify GABAergic projection neurons. GABAergic projection neurons account for 0.5% of the neocortical GABAergic neurons. To elucidate the role of the GABAergic projection neurons in the neocortex, it will be necessary to clarify the network constructed by nNOS-positive GABAergic projection neurons and their postsynaptic targets. Thus, our long-range goals will be to label and manipulate (including deleting) the GABAergic projection neurons using genetic tools driven by a nNOS promoter. We recognize that this may be a complex endeavor, as most excitatory neurons in the murine neocortex express nNOS transiently. Nevertheless, additional studies characterizing long-range GABAergic projection neurons will have great value to the overall understanding of mature cortical function.
Keywords: GABA; GABAergic projection neuron; gamma-oscillations; nNOS; neocortex.
Figures
References
-
- Bayer S. A., Altman J. (1991). Neocortical Development. New York: Raven Press
LinkOut - more resources
Full Text Sources