

Structure of a classical MHC class I molecule that binds "non-classical" ligands
- PMID: 21151886
- PMCID: PMC2998441
- DOI: 10.1371/journal.pbio.1000557
Structure of a classical MHC class I molecule that binds "non-classical" ligands
Abstract
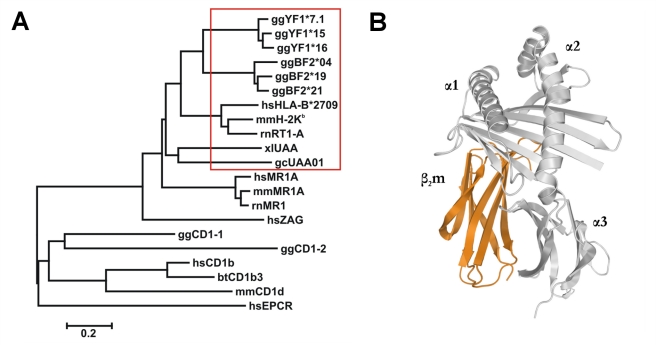
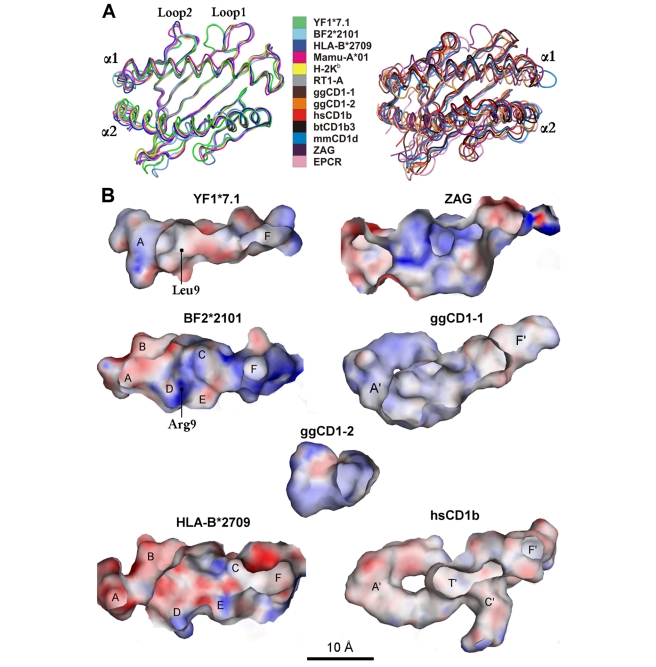
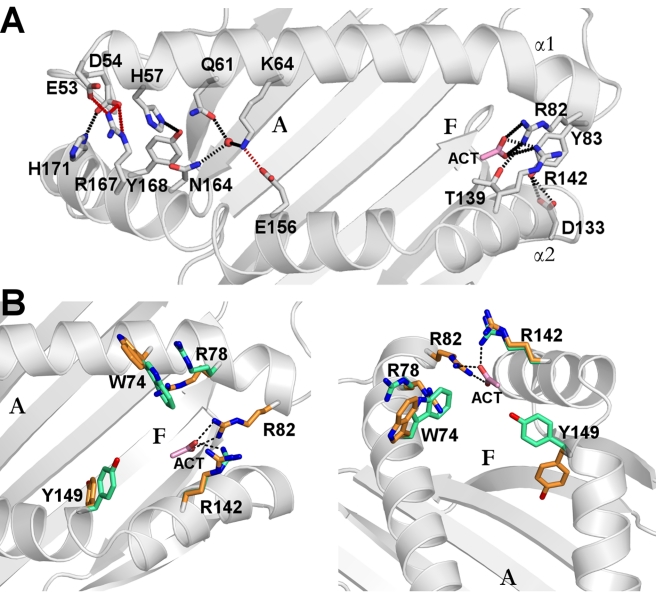
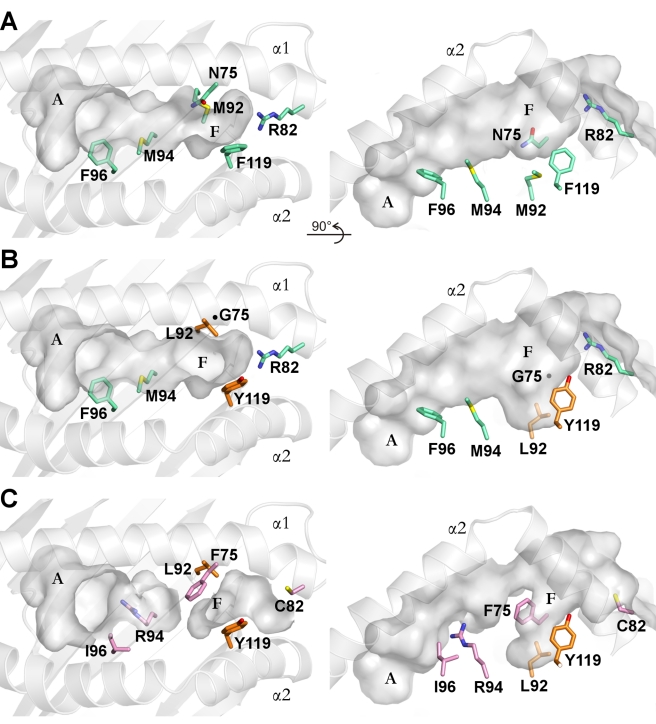
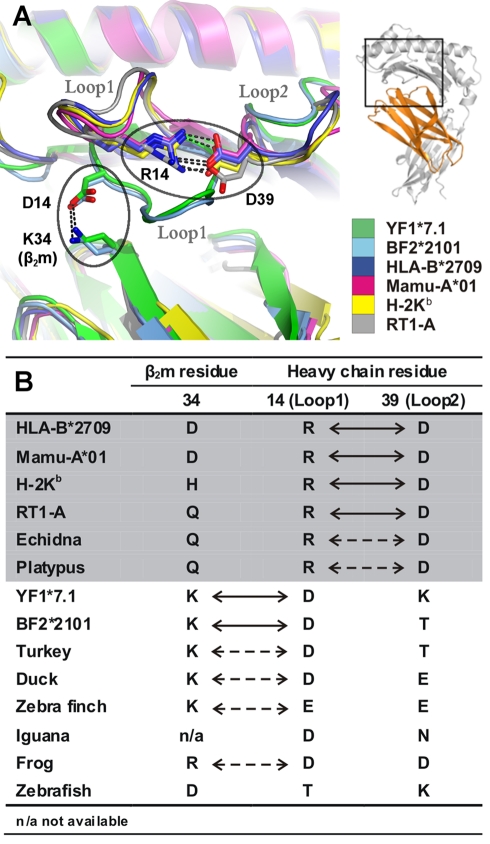
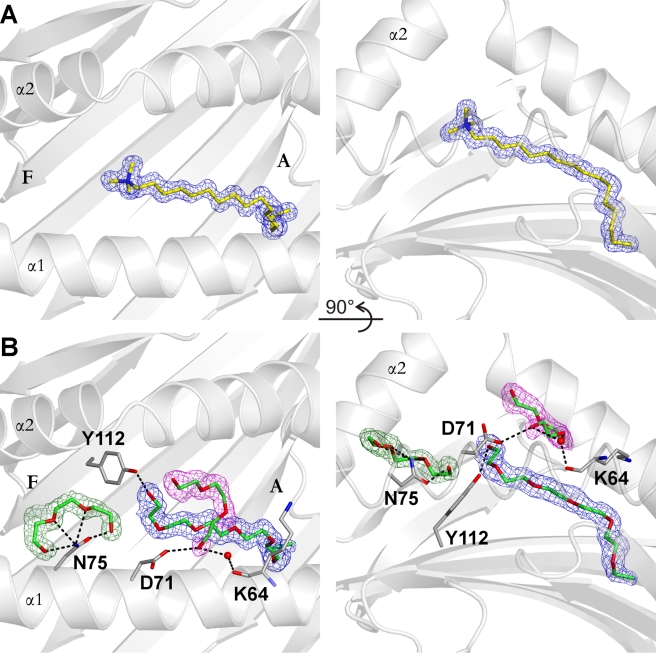
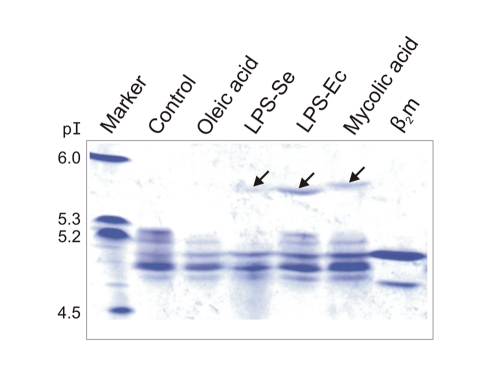
Chicken YF1 genes share a close sequence relationship with classical MHC class I loci but map outside of the core MHC region. To obtain insights into their function, we determined the structure of the YF1*7.1/β(2)-microgloblin complex by X-ray crystallography at 1.3 Å resolution. It exhibits the architecture typical of classical MHC class I molecules but possesses a hydrophobic binding groove that contains a non-peptidic ligand. This finding prompted us to reconstitute YF1*7.1 also with various self-lipids. Seven additional YF1*7.1 structures were solved, but only polyethyleneglycol molecules could be modeled into the electron density within the binding groove. However, an assessment of YF1*7.1 by native isoelectric focusing indicated that the molecules were also able to bind nonself-lipids. The ability of YF1*7.1 to interact with hydrophobic ligands is unprecedented among classical MHC class I proteins and might aid the chicken immune system to recognize a diverse ligand repertoire with a minimal number of MHC class I molecules.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Pink J. R. L, Koch C, Ziegler A. Immuno-ornithological conversation. In: Steinberg C. M, Lefkovits I, editors. Festschrift in honor of Niels Kaj Jerne on the occasion of his 70th birthday: the immune system. Basel: Karger; 1981. pp. 69–75.
-
- Kelley J, Walter L, Trowsdale J. Comparative genomics of major histocompatibility complexes. Immunogenetics. 2005;56:683–695. - PubMed
-
- Madden D. R. The 3-dimensional structure of peptide-MHC complexes. Ann Rev Immunol. 1995;13:587–622. - PubMed
-
- Silk J. D, Salio M, Brown J, Jones E. Y, Cerundolo V. Structural and functional aspects of lipid binding by CD1 molecules. Ann Rev Cell Dev Biol. 2008;24:369–395. - PubMed
-
- Bennett M. J, Lebron J. A, Bjorkman P. J. Crystal structure of the hereditary haemochromatosis protein HFE complexed with transferrin receptor. Nature. 2000;403:46–53. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
