

LaeA control of velvet family regulatory proteins for light-dependent development and fungal cell-type specificity
- PMID: 21152013
- PMCID: PMC2996326
- DOI: 10.1371/journal.pgen.1001226
LaeA control of velvet family regulatory proteins for light-dependent development and fungal cell-type specificity
Abstract
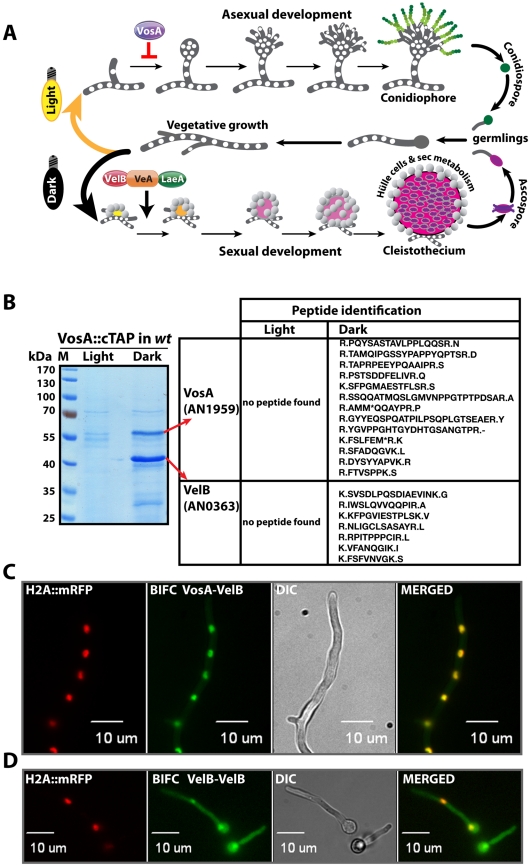
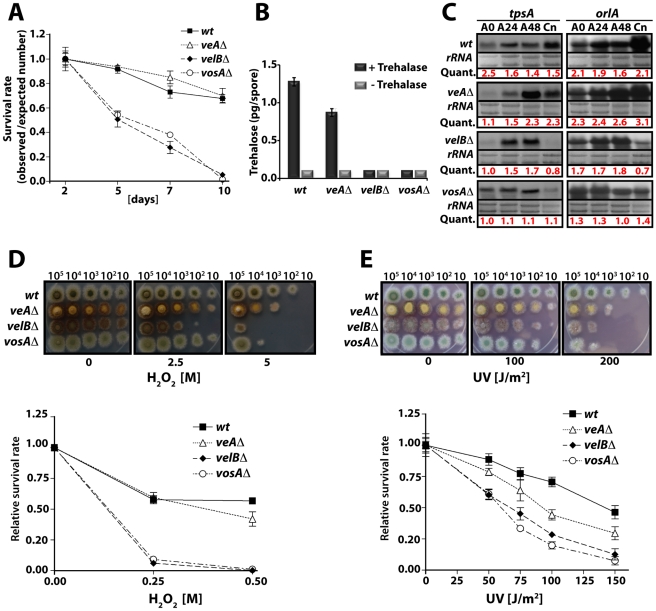
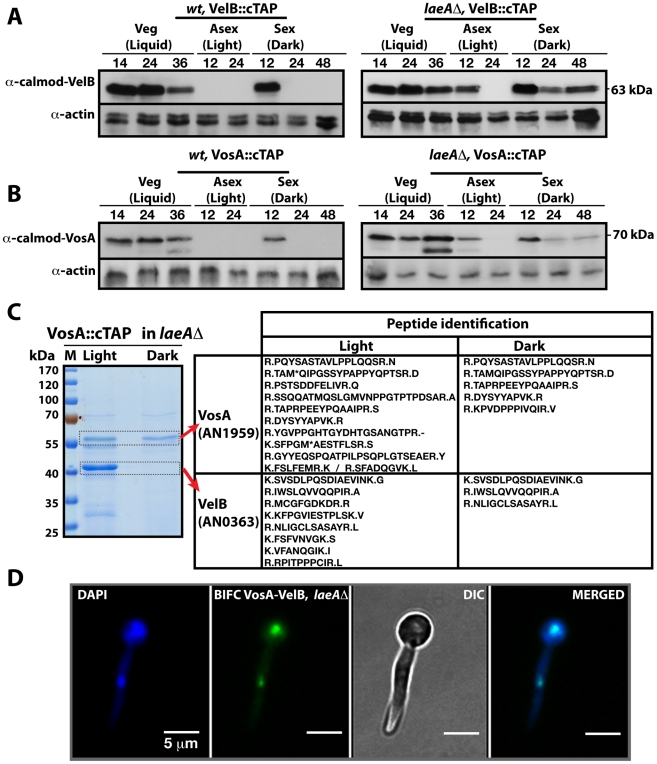
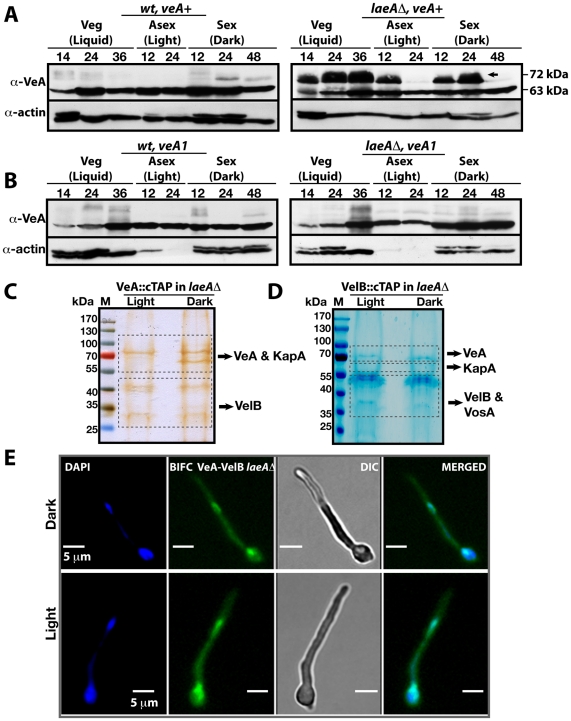
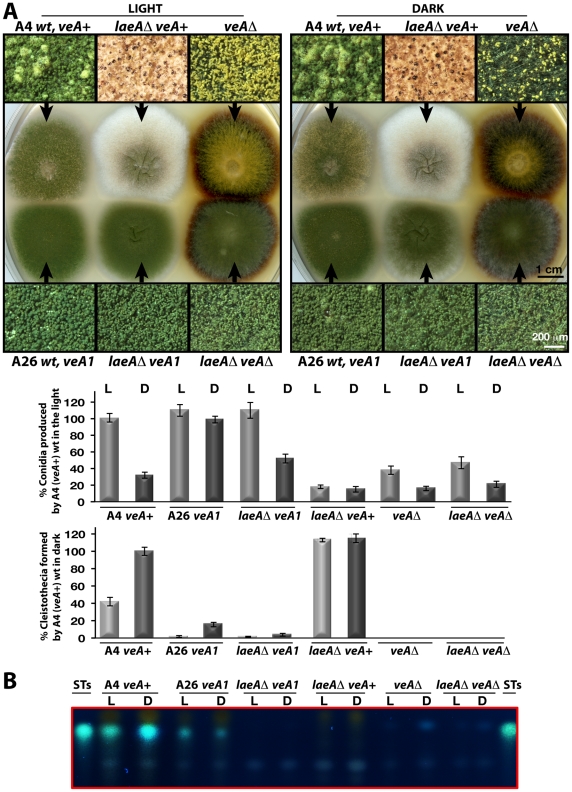
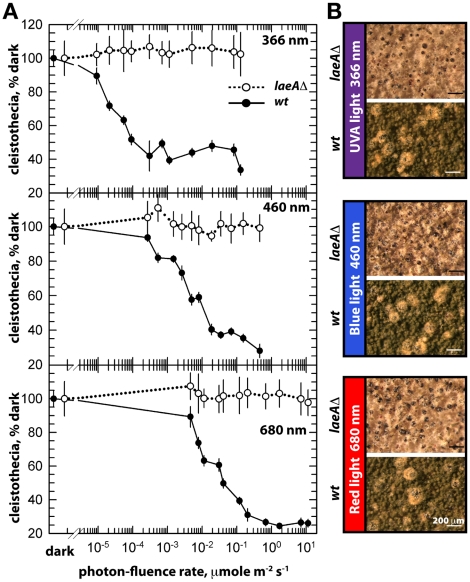
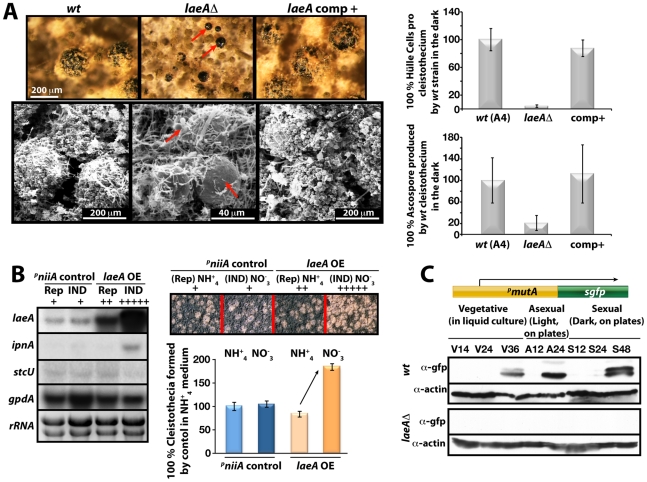
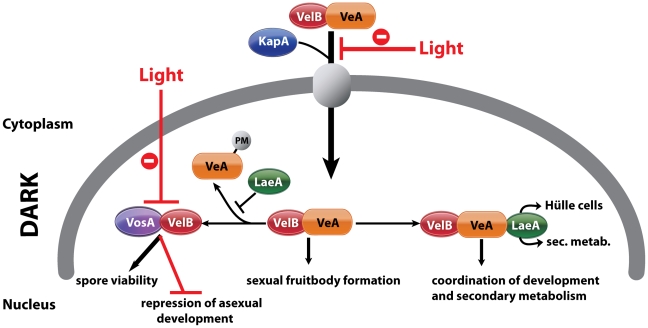
VeA is the founding member of the velvet superfamily of fungal regulatory proteins. This protein is involved in light response and coordinates sexual reproduction and secondary metabolism in Aspergillus nidulans. In the dark, VeA bridges VelB and LaeA to form the VelB-VeA-LaeA (velvet) complex. The VeA-like protein VelB is another developmental regulator, and LaeA has been known as global regulator of secondary metabolism. In this study, we show that VelB forms a second light-regulated developmental complex together with VosA, another member of the velvet family, which represses asexual development. LaeA plays a key role, not only in secondary metabolism, but also in directing formation of the VelB-VosA and VelB-VeA-LaeA complexes. LaeA controls VeA modification and protein levels and possesses additional developmental functions. The laeA null mutant results in constitutive sexual differentiation, indicating that LaeA plays a pivotal role in inhibiting sexual development in response to light. Moreover, the absence of LaeA results in the formation of significantly smaller fruiting bodies. This is due to the lack of a specific globose cell type (Hülle cells), which nurse the young fruiting body during development. This suggests that LaeA controls Hülle cells. In summary, LaeA plays a dynamic role in fungal morphological and chemical development, and it controls expression, interactions, and modification of the velvet regulators.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism.Science. 2008 Jun 13;320(5882):1504-6. doi: 10.1126/science.1155888. Science. 2008. PMID: 18556559
-
Control of Development, Secondary Metabolism and Light-Dependent Carotenoid Biosynthesis by the Velvet Complex of Neurospora crassa.Genetics. 2019 Jul;212(3):691-710. doi: 10.1534/genetics.119.302277. Epub 2019 May 8. Genetics. 2019. PMID: 31068340 Free PMC article.
-
Aspergillus flavus VelB acts distinctly from VeA in conidiation and may coordinate with FluG to modulate sclerotial production.Fungal Genet Biol. 2013 Sep-Oct;58-59:71-9. doi: 10.1016/j.fgb.2013.08.009. Epub 2013 Aug 29. Fungal Genet Biol. 2013. PMID: 23994319
-
[Progress in the study of Velvet and LaeA proteins and their relation to the development and bioactive compounds in medicinal fungi].Yao Xue Xue Bao. 2014 Nov;49(11):1520-7. Yao Xue Xue Bao. 2014. PMID: 25757276 Review. Chinese.
-
Involvement of LaeA and Velvet Proteins in Regulating the Production of Mycotoxins and Other Fungal Secondary Metabolites.J Fungi (Basel). 2024 Aug 8;10(8):561. doi: 10.3390/jof10080561. J Fungi (Basel). 2024. PMID: 39194887 Free PMC article. Review.
Cited by
-
The function of a conidia specific transcription factor CsgA in Aspergillus nidulans.Sci Rep. 2022 Sep 16;12(1):15588. doi: 10.1038/s41598-022-19749-6. Sci Rep. 2022. PMID: 36114253 Free PMC article.
-
Hülle Cells of Aspergillus nidulans with Nuclear Storage and Developmental Backup Functions Are Reminiscent of Multipotent Stem Cells.mBio. 2020 Aug 11;11(4):e01673-20. doi: 10.1128/mBio.01673-20. mBio. 2020. PMID: 32788382 Free PMC article.
-
NsdC and NsdD affect Aspergillus flavus morphogenesis and aflatoxin production.Eukaryot Cell. 2012 Sep;11(9):1104-11. doi: 10.1128/EC.00069-12. Epub 2012 Jul 13. Eukaryot Cell. 2012. PMID: 22798394 Free PMC article.
-
The chromatin code of fungal secondary metabolite gene clusters.Appl Microbiol Biotechnol. 2012 Sep;95(6):1389-404. doi: 10.1007/s00253-012-4208-8. Epub 2012 Jul 20. Appl Microbiol Biotechnol. 2012. PMID: 22814413 Free PMC article. Review.
-
Construction of Light-Responsive Gene Regulatory Network for Growth, Development and Secondary Metabolite Production in Cordyceps militaris.Biology (Basel). 2022 Jan 4;11(1):71. doi: 10.3390/biology11010071. Biology (Basel). 2022. PMID: 35053069 Free PMC article.
References
-
- Huse JT, Holland EC. Targeting brain cancer: advances in the molecular pathology of malignant glioma and medulloblastoma. Nat Rev Cancer. 2010;10:319–331. - PubMed
-
- Braus GH, Krappmann S, Eckert SE. Sexual Development in Ascomycetes - Fruit Body Formation of Aspergillus nidulans. In: Osiewacz HD, editor. Molecular Biology of Fungal Development. New York, Basel: Marcel Dekker, Inc; 2002. pp. 215–244.
-
- Pöggeler S, Nowrousian M, Kück U. Fruiting-Body Development in Ascomycetes. In: Fischer K, editor. The Mycota I Growth, Differentiation and Sexuality. Heidelberg: Springer-Verlag; 2006. pp. 325–355.
-
- Sohn KT, Yoon KS. Ultrastructural Study on the Cleistothecium Development in Aspergillus nidulans. Mycobiology. 2002;30:117–127.
-
- Keller NP, Turner G, Bennett JW. Fungal secondary metabolism - from biochemistry to genomics. Nat Rev Microbiol. 2005;3:937–947. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
