

Model for Protein Concentration Gradients in the Cytoplasm
- PMID: 21152415
- PMCID: PMC2996619
- DOI: 10.1007/s12195-008-0008-8
Model for Protein Concentration Gradients in the Cytoplasm
Abstract
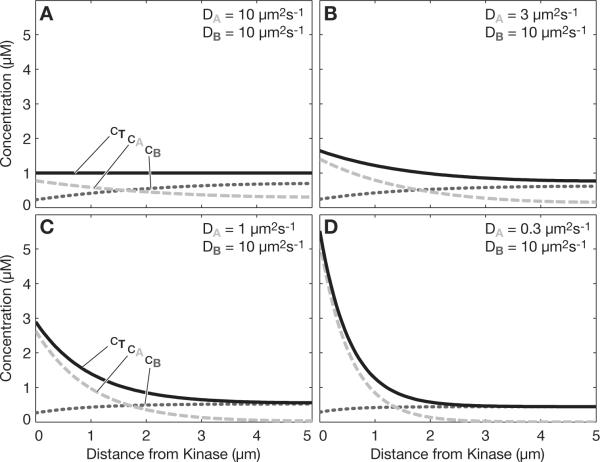
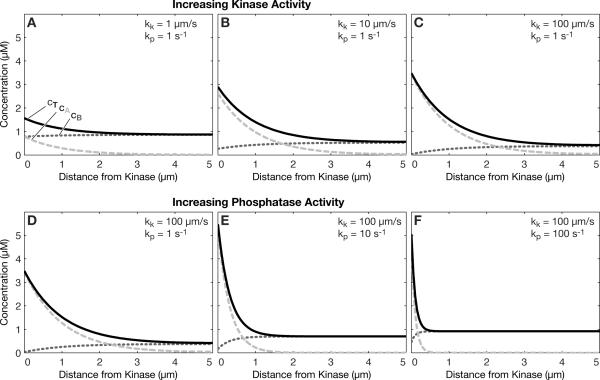
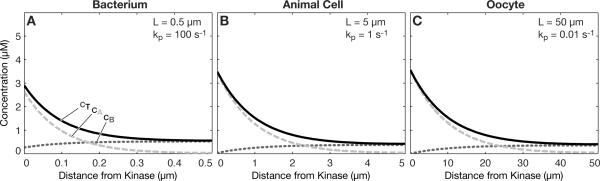
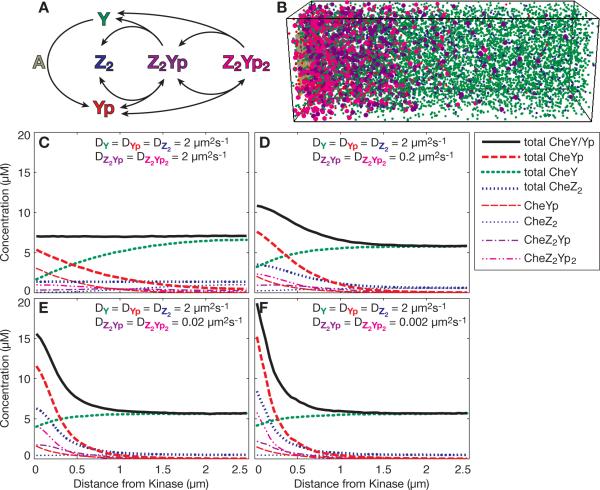
Intracellular protein concentration gradients are generally thought to be unsustainable at steady-state due to diffusion. Here we show how protein concentration gradients can theoretically be sustained indefinitely through a relatively simple mechanism that couples diffusion to a spatially segregated kinase-phosphatase system. Although it is appreciated that such systems can theoretically give rise to phosphostate gradients, it has been assumed that they do not give rise to gradients in the total protein concentration. Here we show that this assumption does not hold if the two forms of protein have different diffusion coefficients. If, for example, the phosphorylated state binds selectively to a second larger protein or protein complex then a steady state gradient in total protein concentration will be created. We illustrate the principle with an analytical solution to the diffusion-reaction problem and by stochastic individual-based simulations using the Smoldyn program. We argue that protein gradients created in this way need to be considered in experiments using fluorescent probes and could in principle encode spatial information in the cytoplasm.
Figures
References
-
- Andrews SS, Bray D. Stochastic simulation of chemical reactions with spatial resolution and single molecule detail. Phys Biol. 2004;1:137–51. - PubMed
-
- Blat Y, Eisenbach M. Phosphorylation-dependent binding of the chemotaxis signal molecule CheY to its phosphatase, CheZ. Biochemistry. 1994;33(4):902–6. - PubMed
-
- Brown GC, Kholodenko BN. Spatial gradients of cellular phospho-proteins. FEBS Lett. 1999;457(3):452–4. - PubMed
-
- Cassimeris L. The oncoprotein 18/stathmin family of microtubule destabilizers. Curr Opin Cell Biol. 2002;14(1):18–24. - PubMed
-
- Caudron M, Bunt G, Bastiaens P, Karsenti E. Spatial coordination of spindle assembly by chromosome-mediated signaling gradients. Science. 2005;309(5739):1373–6. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
