

The function and three-dimensional structure of a thromboxane A2/cysteinyl leukotriene-binding protein from the saliva of a mosquito vector of the malaria parasite
- PMID: 21152418
- PMCID: PMC2994686
- DOI: 10.1371/journal.pbio.1000547
The function and three-dimensional structure of a thromboxane A2/cysteinyl leukotriene-binding protein from the saliva of a mosquito vector of the malaria parasite
Abstract
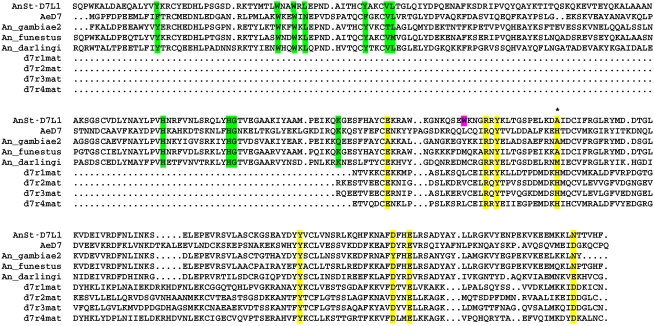
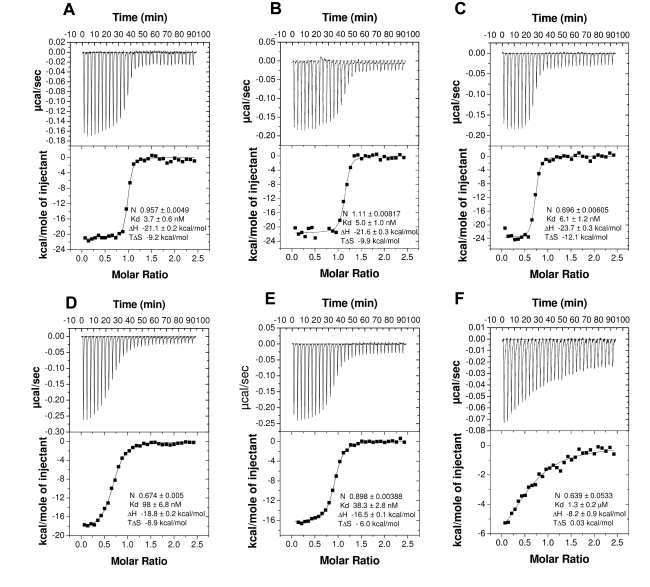
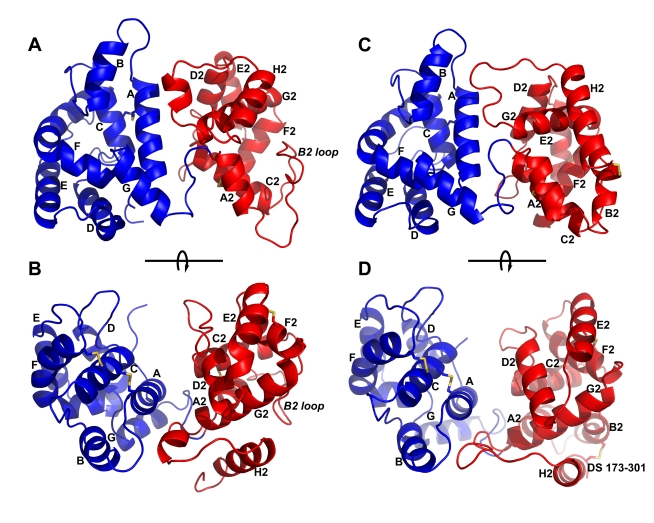
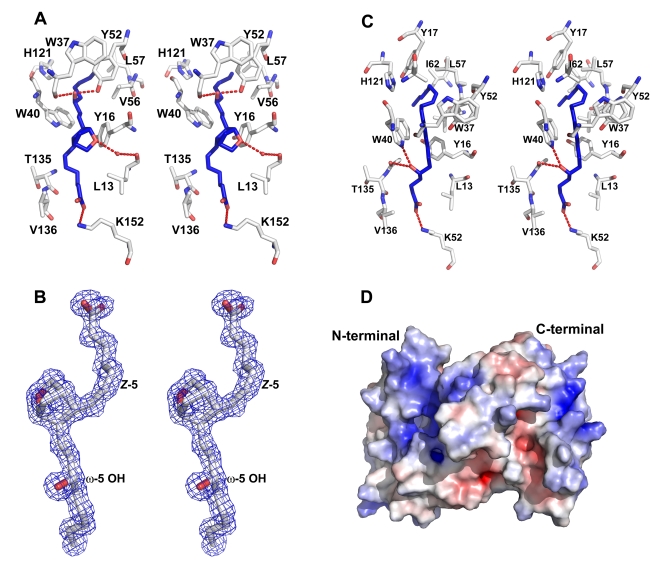
The highly expressed D7 protein family of mosquito saliva has previously been shown to act as an anti-inflammatory mediator by binding host biogenic amines and cysteinyl leukotrienes (CysLTs). In this study we demonstrate that AnSt-D7L1, a two-domain member of this group from Anopheles stephensi, retains the CysLT binding function seen in the homolog AeD7 from Aedes aegypti but has lost the ability to bind biogenic amines. Unlike any previously characterized members of the D7 family, AnSt-D7L1 has acquired the important function of binding thromboxane A(2) (TXA(2)) and its analogs with high affinity. When administered to tissue preparations, AnSt-D7L1 abrogated Leukotriene C(4) (LTC(4))-induced contraction of guinea pig ileum and contraction of rat aorta by the TXA(2) analog U46619. The protein also inhibited platelet aggregation induced by both collagen and U46619 when administered to stirred platelets. The crystal structure of AnSt-D7L1 contains two OBP-like domains and has a structure similar to AeD7. In AnSt-D7L1, the binding pocket of the C-terminal domain has been rearranged relative to AeD7, making the protein unable to bind biogenic amines. Structures of the ligand complexes show that CysLTs and TXA(2) analogs both bind in the same hydrophobic pocket of the N-terminal domain. The TXA(2) analog U46619 is stabilized by hydrogen bonding interactions of the ω-5 hydroxyl group with the phenolic hydroxyl group of Tyr 52. LTC(4) and occupies a very similar position to LTE(4) in the previously determined structure of its complex with AeD7. As yet, it is not known what, if any, new function has been acquired by the rearranged C-terminal domain. This article presents, to our knowledge, the first structural characterization of a protein from mosquito saliva that inhibits collagen mediated platelet activation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





References
-
- Ribeiro J. M. Blood-feeding arthropods: live syringes or invertebrate pharmacologists? Infect Agents Dis. 1995;4:143–152. - PubMed
-
- Ribeiro J. M, Francischetti I. M. Role of arthropod saliva in blood feeding: sialome and post-sialome perspectives. Annu Rev Entomol. 2003;48:73–88. - PubMed
-
- Champagne D. E. Antihemostatic molecules from saliva of blood-feeding arthropods. Pathophysiol Haemost Thromb. 2005;34:221–227. - PubMed
-
- Valenzuela J. G. High-throughput approaches to study salivary proteins and genes from vectors of disease. Insect Biochem Mol Biol. 2002;32:1199–1209. - PubMed
-
- Titus R. G, Ribeiro J. M. Salivary gland lysates from the sand fly Lutzomyia longipalpis enhance Leishmania infectivity. Science. 1988;239:1306–1308. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
