

Three groups of interneurons account for nearly 100% of neocortical GABAergic neurons
- PMID: 21154909
- PMCID: PMC3556905
- DOI: 10.1002/dneu.20853
Three groups of interneurons account for nearly 100% of neocortical GABAergic neurons
Abstract
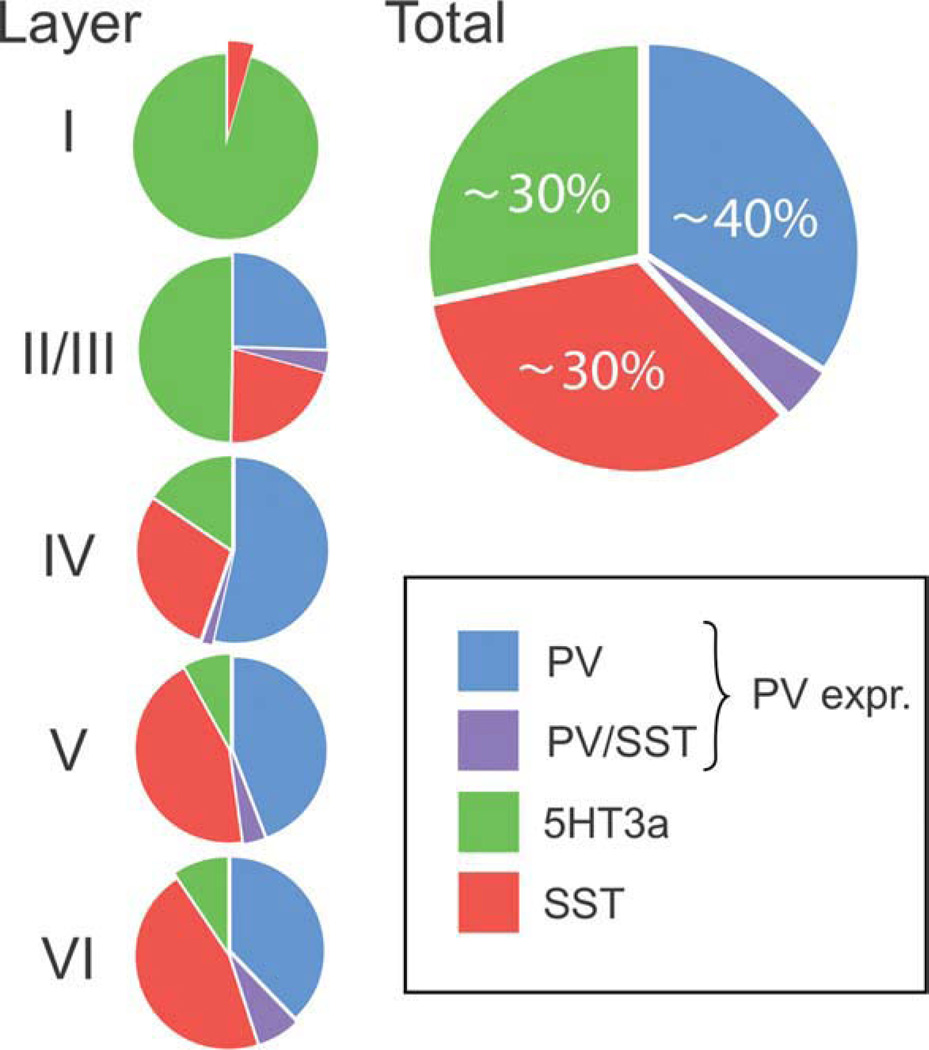
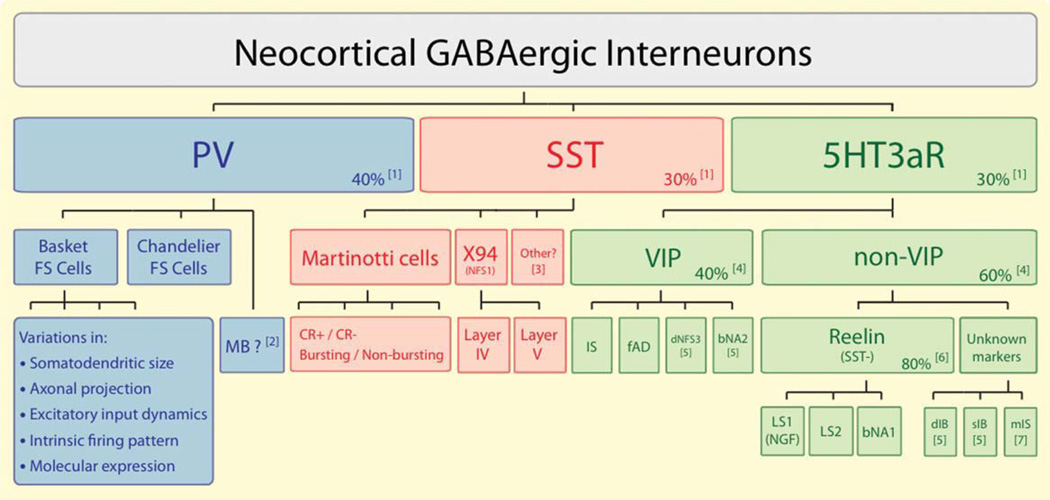
An understanding of the diversity of cortical GABAergic interneurons is critical to understand the function of the cerebral cortex. Recent data suggest that neurons expressing three markers, the Ca2+-binding protein parvalbumin (PV), the neuropeptide somatostatin (SST), and the ionotropic serotonin receptor 5HT3a (5HT3aR) account for nearly 100% of neocortical interneurons. Interneurons expressing each of these markers have a different embryological origin. Each group includes several types of interneurons that differ in morphological and electrophysiological properties and likely have different functions in the cortical circuit. The PV group accounts for ∼40% of GABAergic neurons and includes fast spiking basket cells and chandelier cells. The SST group, which represents ∼30% of GABAergic neurons, includes the Martinotti cells and a set of neurons that specifically target layerIV. The 5HT3aR group, which also accounts for ∼30% of the total interneuronal population, is heterogeneous and includes all of the neurons that express the neuropeptide VIP, as well as an equally numerous subgroup of neurons that do not express VIP and includes neurogliaform cells. The universal modulation of these neurons by serotonin and acetylcholine via ionotropic receptors suggests that they might be involved in shaping cortical circuits during specific brain states and behavioral contexts.
Copyright © 2010 Wiley Periodicals, Inc.
Figures
References
-
- Acsady L, Gorcs TJ, Freund TF. Different populations of vasoactive intestinal polypeptide-immunoreactive interneurons are specialized to control pyramidal cells or interneurons in the hippocampus. Neuroscience. 1996;73:317–334. - PubMed
-
- Anderson SA, Eisenstat DD, Shi L, Rubenstein JL. Interneuron migration from basal forebrain to neocortex: Dependence on Dlx genes. Science. 1997;278:474–476. - PubMed
-
- Bacci A, Huguenard JR, Prince DA. Modulation of neocortical interneurons: Extrinsic influences and exercises in self-control. Trends Neurosci. 2005;28:602–610. - PubMed
-
- Bartos M, Vida I, Jonas P. Synaptic mechanisms of synchronized gamma oscillations in inhibitory interneuron networks. Nat Rev. 2007;8:45–56. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
