

Trichoderma viride cellulase induces resistance to the antibiotic pore-forming peptide alamethicin associated with changes in the plasma membrane lipid composition of tobacco BY-2 cells
- PMID: 21156059
- PMCID: PMC3017840
- DOI: 10.1186/1471-2229-10-274
Trichoderma viride cellulase induces resistance to the antibiotic pore-forming peptide alamethicin associated with changes in the plasma membrane lipid composition of tobacco BY-2 cells
Abstract
Background: Alamethicin is a membrane-active peptide isolated from the beneficial root-colonising fungus Trichoderma viride. This peptide can insert into membranes to form voltage-dependent pores. We have previously shown that alamethicin efficiently permeabilises the plasma membrane, mitochondria and plastids of cultured plant cells. In the present investigation, tobacco cells (Nicotiana tabacum L. cv Bright Yellow-2) were pre-treated with elicitors of defence responses to study whether this would affect permeabilisation.
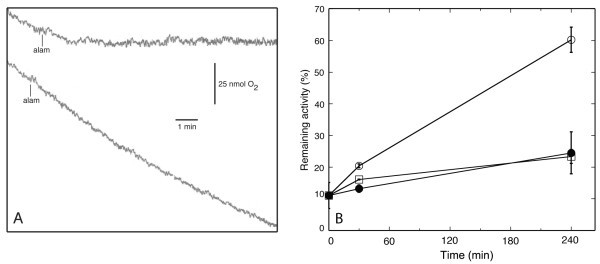
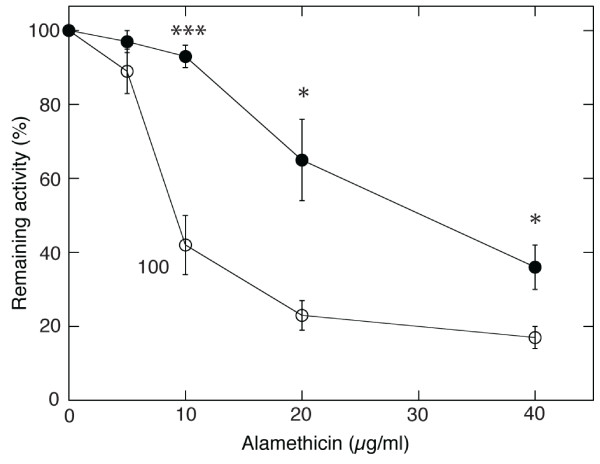
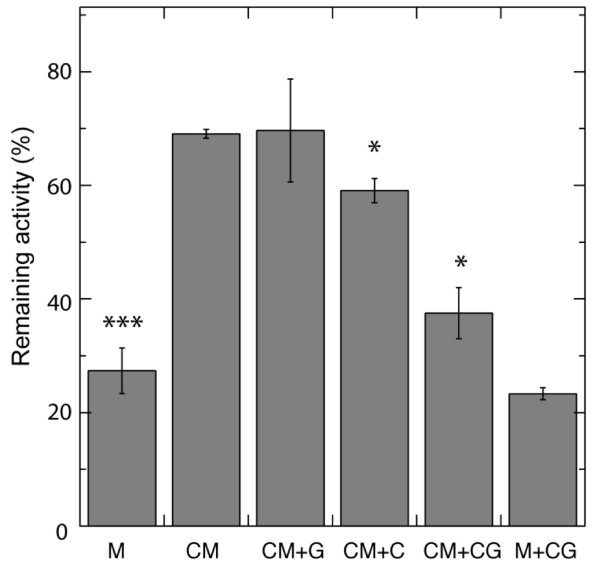
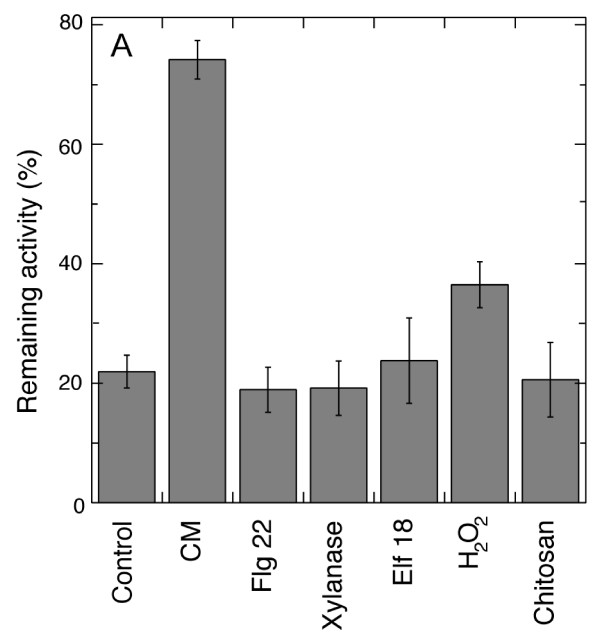
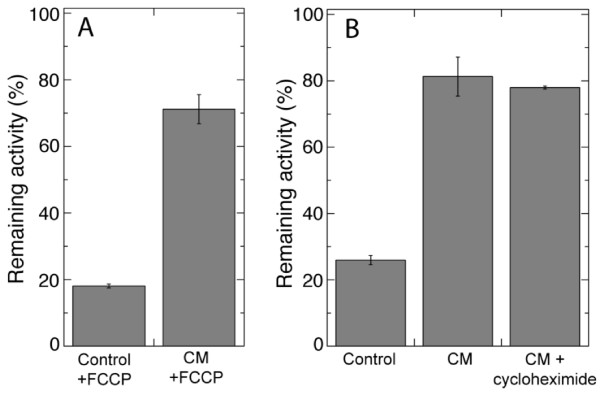
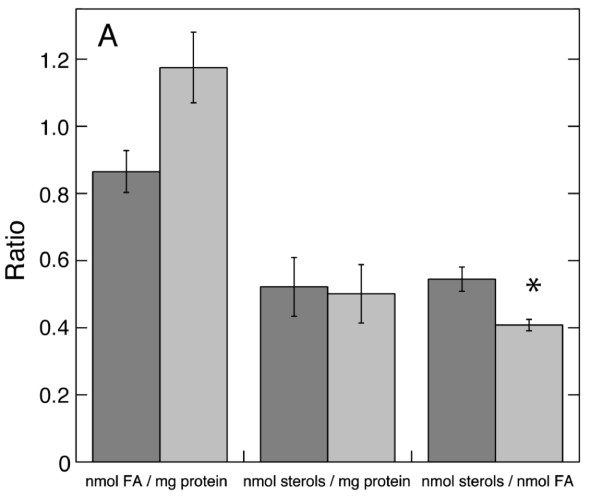
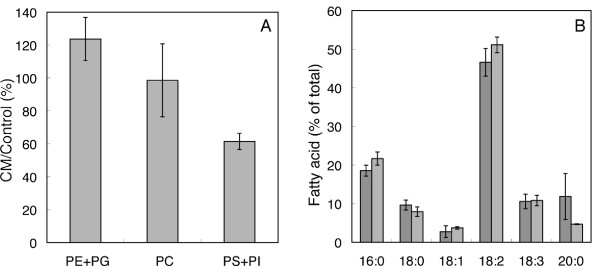
Results: Oxygen consumption experiments showed that added cellulase, already upon a limited cell wall digestion, induced a cellular resistance to alamethicin permeabilisation. This effect could not be elicited by xylanase or bacterial elicitors such as flg22 or elf18. The induction of alamethicin resistance was independent of novel protein synthesis. Also, the permeabilisation was unaffected by the membrane-depolarising agent FCCP. As judged by lipid analyses, isolated plasma membranes from cellulase-pretreated tobacco cells contained less negatively charged phospholipids (PS and PI), yet higher ratios of membrane lipid fatty acid to sterol and to protein, as compared to control membranes.
Conclusion: We suggest that altered membrane lipid composition as induced by cellulase activity may render the cells resistant to alamethicin. This induced resistance could reflect a natural process where the plant cells alter their sensitivity to membrane pore-forming agents secreted by Trichoderma spp. to attack other microorganisms, and thus adding to the beneficial effect that Trichoderma has for plant root growth. Furthermore, our data extends previous reports on artificial membranes on the importance of lipid packing and charge for alamethicin permeabilisation to in vivo conditions.
Figures
References
-
- Keen NT, Tamaki S, Kobayashi D, Gerhold D, Stayton M, Shen H, Gold S, Lorang J, Thordal-Christensen H, Dahlbeck D. et al.Bacteria expressing avirulence Gene D produce a specific elicitor of the soybean hypersensitive reaction. Mol Plant Microbe Interact. 1990;3:112–121.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
