

Local geometry and elasticity in compact chromatin structure
- PMID: 21156136
- PMCID: PMC3000514
- DOI: 10.1016/j.bpj.2010.10.024
Local geometry and elasticity in compact chromatin structure
Abstract
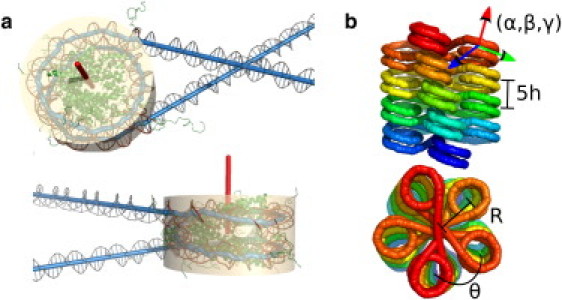
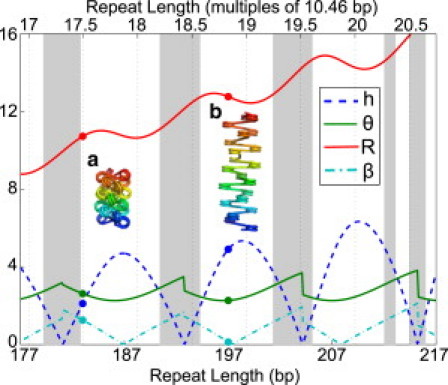
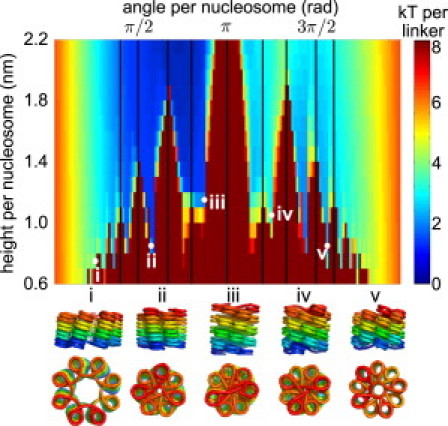
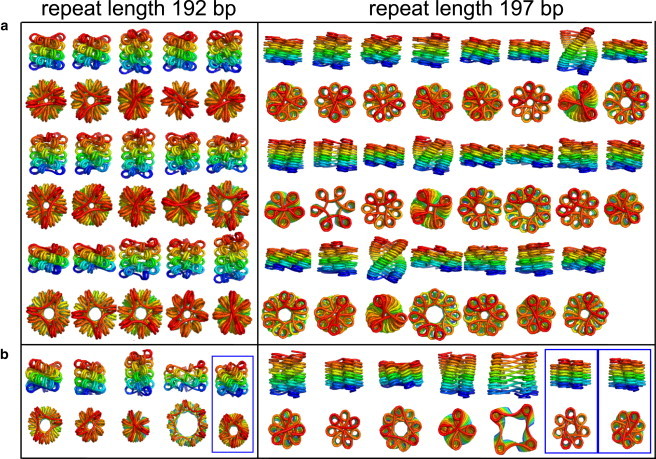
The hierarchical packaging of DNA into chromatin within a eukaryotic nucleus plays a pivotal role in both the accessibility of genomic information and the dynamics of replication. Our work addresses the role of nanoscale physical and geometric properties in determining the structure of chromatin at the mesoscale level. We study the packaging of DNA in chromatin fibers by optimization of regular helical morphologies, considering the elasticity of the linker DNA as well as steric packing of the nucleosomes and linkers. Our model predicts a broad range of preferred helix structures for a fixed linker length of DNA; changing the linker length alters the predicted ensemble. Specifically, we find that the twist registry of the nucleosomes, as set by the internucleosome repeat length, determines the preferred angle between the nucleosomes and the fiber axis. For moderate to long linker lengths, we find a number of energetically comparable configurations with different nucleosome-nucleosome interaction patterns, indicating a potential role for kinetic trapping in chromatin fiber formation. Our results highlight the key role played by DNA elasticity and local geometry in regulating the hierarchical packaging of the genome.
Copyright © 2010 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Richmond T.J., Davey C.A. The structure of DNA in the nucleosome core. Nature. 2003;423:145–150. - PubMed
-
- Huynh V.A., Robinson P.J., Rhodes D. A method for the in vitro reconstitution of a defined “30 nm” chromatin fiber containing stoichiometric amounts of the linker histone. J. Mol. Biol. 2005;345:957–968. - PubMed
-
- Langmore J.P., Schutt C. The higher order structure of chicken erythrocyte chromosomes in vivo. Nature. 1980;288:620–622. - PubMed
-
- Kruithof M., Chien F.T., van Noort J. Single-molecule force spectroscopy reveals a highly compliant helical folding for the 30-nm chromatin fiber. Nat. Struct. Mol. Biol. 2009;16:534–540. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
