

Effect of phospholipidosis on the cellular pharmacokinetics of chloroquine
- PMID: 21156819
- PMCID: PMC3061524
- DOI: 10.1124/jpet.110.175679
Effect of phospholipidosis on the cellular pharmacokinetics of chloroquine
Abstract
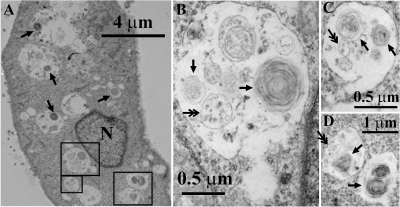
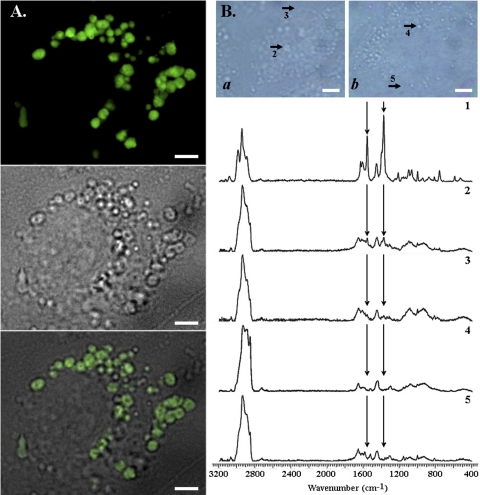
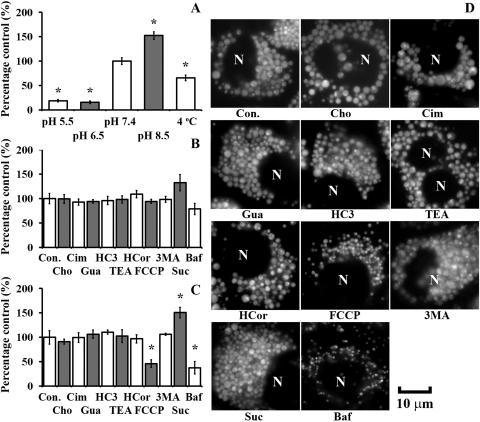
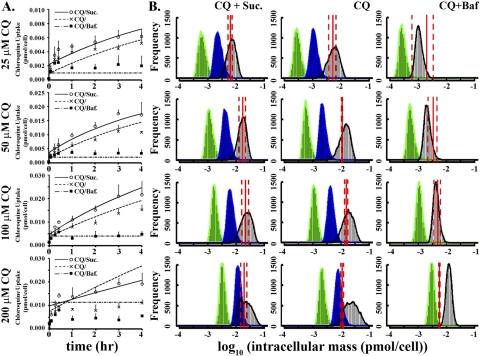
In vivo, the weakly basic, lipophilic drug chloroquine (CQ) accumulates in the kidney to concentrations more than a thousand-fold greater than those in plasma. To study the cellular pharmacokinetics of chloroquine in cells derived from the distal tubule, Madin-Darby canine kidney cells were incubated with CQ under various conditions. CQ progressively accumulated without exhibiting steady-state behavior. Experiments failed to yield evidence that known active transport mechanisms mediated CQ uptake at the plasma membrane. CQ induced a phospholipidosis-like phenotype, characterized by the appearance of numerous multivesicular and multilamellar bodies (MLBs/MVBs) within the lumen of expanded cytoplasmic vesicles. Other induced phenotypic changes including changes in the volume and pH of acidic organelles were measured, and the integrated effects of all these changes were computationally modeled to establish their impact on intracellular CQ mass accumulation. Based on the passive transport behavior of CQ, the measured phenotypic changes fully accounted for the continuous, nonsteady-state CQ accumulation kinetics. Consistent with the simulation results, Raman confocal microscopy of live cells confirmed that CQ became highly concentrated within induced, expanded cytoplasmic vesicles that contained multiple MLBs/MVBs. Progressive CQ accumulation was increased by sucrose, a compound that stimulated the phospholipidosis-like phenotype, and was decreased by bafilomycin A1, a compound that inhibited this phenotype. Thus, phospholipidosis-associated changes in organelle structure and intracellular membrane content can exert a major influence on the local bioaccumulation and biodistribution of drugs.
Figures

References
-
- Bawolak MT, Morissette G, Marceau F. (2010) Vacuolar ATPase-mediated sequestration of local anesthetics in swollen macroautophagosomes. Can J Anaesth 57:230–239 - PubMed
-
- de Duve C, de Barsy T, Poole B, Trouet A, Tulkens P, Van Hoof F. (1974) Commentary. Lysosomotropic agents. Biochem Pharmacol 23:2495–2531 - PubMed
-
- Djordevic B, Lange CS, Rotman M. (1992) Potentiation of radiation lethality in mouse melanoma cells by mild hyperthermia and chloroquine. Melanoma Res 2:321–326 - PubMed
-
- Ducharme J, Farinotti R. (1996) Clinical pharmacokinetics and metabolism of chloroquine. Focus on recent advancements. Clin Pharmacokinet 31:257–274 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
