

SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation
- PMID: 21156857
- PMCID: PMC3032448
- DOI: 10.1104/pp.110.165191
SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation
Abstract
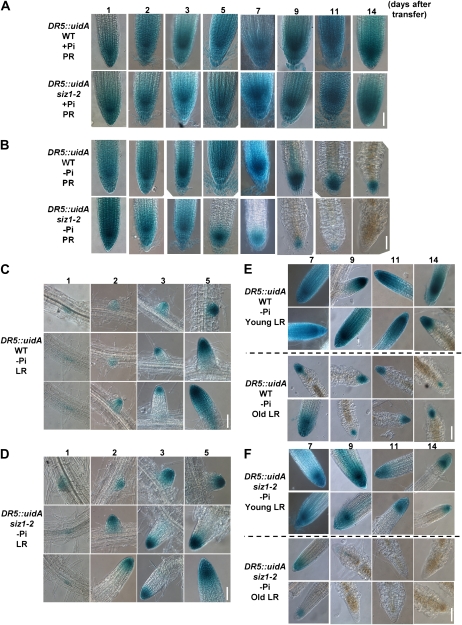
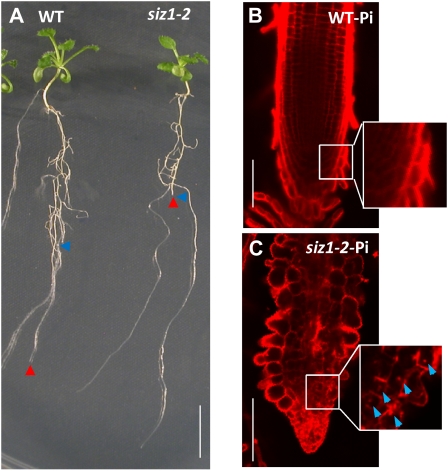
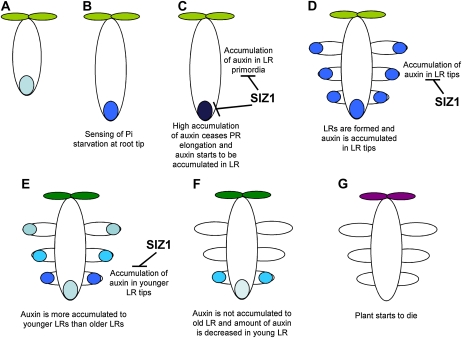
Phosphate (Pi) limitation causes plants to modulate the architecture of their root systems to facilitate the acquisition of Pi. Previously, we reported that the Arabidopsis (Arabidopsis thaliana) SUMO E3 ligase SIZ1 regulates root architecture remodeling in response to Pi limitation; namely, the siz1 mutations cause the inhibition of primary root (PR) elongation and the promotion of lateral root (LR) formation. Here, we present evidence that SIZ1 is involved in the negative regulation of auxin patterning to modulate root system architecture in response to Pi starvation. The siz1 mutations caused greater PR growth inhibition and LR development of seedlings in response to Pi limitation. Similar root phenotypes occurred if Pi-deficient wild-type seedlings were supplemented with auxin. N-1-Naphthylphthalamic acid, an inhibitor of auxin efflux activity, reduced the Pi starvation-induced LR root formation of siz1 seedlings to a level equivalent to that seen in the wild type. Monitoring of the auxin-responsive reporter DR5::uidA indicated that auxin accumulates in PR tips at early stages of the Pi starvation response. Subsequently, DR5::uidA expression was observed in the LR primordia, which was associated with LR elongation. The time-sequential patterning of DR5::uidA expression occurred earlier in the roots of siz1 as compared with the wild type. In addition, microarray analysis revealed that several other auxin-responsive genes, including genes involved in cell wall loosening and biosynthesis, were up-regulated in siz1 relative to wild-type seedlings in response to Pi starvation. Together, these results suggest that SIZ1 negatively regulates Pi starvation-induced root architecture remodeling through the control of auxin patterning.
Figures







Similar articles
-
The function of LPR1 is controlled by an element in the promoter and is independent of SUMO E3 Ligase SIZ1 in response to low Pi stress in Arabidopsis thaliana.Plant Cell Physiol. 2010 Mar;51(3):380-94. doi: 10.1093/pcp/pcq004. Epub 2010 Jan 12. Plant Cell Physiol. 2010. PMID: 20071375
-
The Arabidopsis SUMO E3 ligase SIZ1 controls phosphate deficiency responses.Proc Natl Acad Sci U S A. 2005 May 24;102(21):7760-5. doi: 10.1073/pnas.0500778102. Epub 2005 May 13. Proc Natl Acad Sci U S A. 2005. PMID: 15894620 Free PMC article.
-
Overexpression of GbWRKY1 positively regulates the Pi starvation response by alteration of auxin sensitivity in Arabidopsis.Plant Cell Rep. 2012 Dec;31(12):2177-88. doi: 10.1007/s00299-012-1328-7. Epub 2012 Aug 14. Plant Cell Rep. 2012. PMID: 22890372
-
Root architecture remodeling induced by phosphate starvation.Plant Signal Behav. 2011 Aug;6(8):1122-6. doi: 10.4161/psb.6.8.15752. Epub 2011 Aug 1. Plant Signal Behav. 2011. PMID: 21778826 Free PMC article. Review.
-
Transcriptional regulation of phosphate acquisition by higher plants.Cell Mol Life Sci. 2012 Oct;69(19):3207-24. doi: 10.1007/s00018-012-1090-6. Epub 2012 Aug 17. Cell Mol Life Sci. 2012. PMID: 22899310 Free PMC article. Review.
Cited by
-
The SUMO E3 Ligase MdSIZ1 Sumoylates a Cell Number Regulator MdCNR8 to Control Organ Size.Front Plant Sci. 2022 Apr 15;13:836935. doi: 10.3389/fpls.2022.836935. eCollection 2022. Front Plant Sci. 2022. PMID: 35498700 Free PMC article.
-
Loci and natural alleles underlying robust roots and adaptive domestication of upland ecotype rice in aerobic conditions.PLoS Genet. 2018 Aug 10;14(8):e1007521. doi: 10.1371/journal.pgen.1007521. eCollection 2018 Aug. PLoS Genet. 2018. PMID: 30096145 Free PMC article.
-
PI4KIIIβ Activity Regulates Lateral Root Formation Driven by Endocytic Trafficking to the Vacuole.Plant Physiol. 2019 Sep;181(1):112-126. doi: 10.1104/pp.19.00695. Epub 2019 Jul 8. Plant Physiol. 2019. PMID: 31285293 Free PMC article.
-
Cold signaling and cold response in plants.Int J Mol Sci. 2013 Mar 6;14(3):5312-37. doi: 10.3390/ijms14035312. Int J Mol Sci. 2013. PMID: 23466881 Free PMC article.
-
Roots Withstanding their Environment: Exploiting Root System Architecture Responses to Abiotic Stress to Improve Crop Tolerance.Front Plant Sci. 2016 Aug 31;7:1335. doi: 10.3389/fpls.2016.01335. eCollection 2016. Front Plant Sci. 2016. PMID: 27630659 Free PMC article. Review.
References
-
- Abel S, Ticconi CA, Delatorre CA. (2002) Phosphate sensing in higher plants. Physiol Plant 115: 1–8 - PubMed
-
- Al-Ghazi Y, Muller B, Pinloche S, Tranbarger TG, Nacry P, Rossignol M, Tardieu F, Doumas P. (2003) Temporal responses of Arabidopsis root architecture to phosphate starvation: evidence for the involvement of auxin signalling. Plant Cell Environ 26: 1053–1066
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
