

Cardiac neural crest orchestrates remodeling and functional maturation of mouse semilunar valves
- PMID: 21157040
- PMCID: PMC3007154
- DOI: 10.1172/JCI44244
Cardiac neural crest orchestrates remodeling and functional maturation of mouse semilunar valves
Abstract
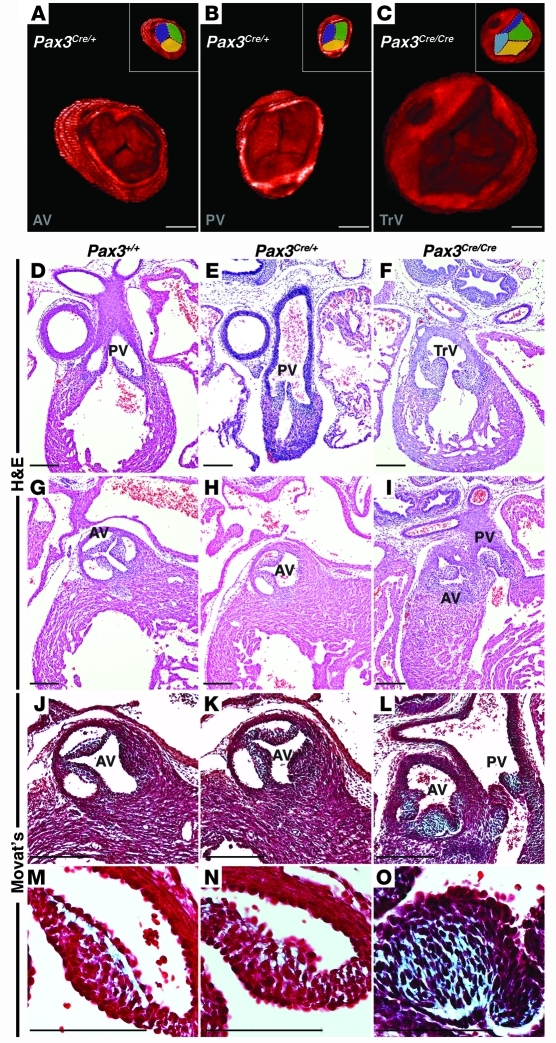
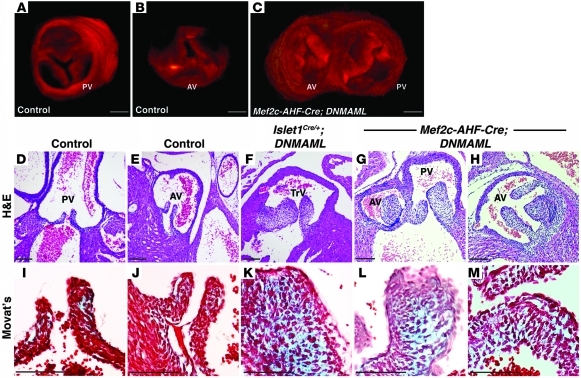
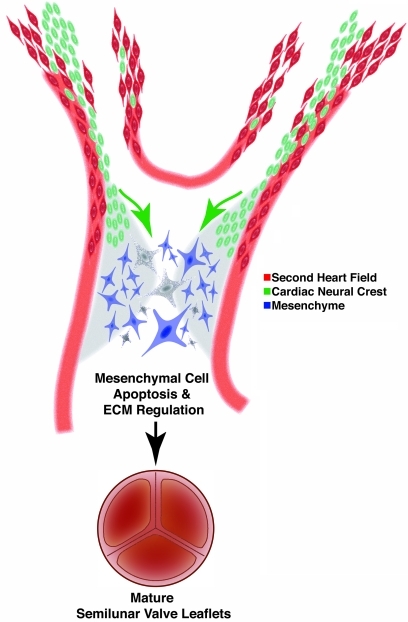
Congenital anomalies of the aortic valve are common and are associated with progressive valvular insufficiency and/or stenosis. In addition, aneurysm, coarctation, and dissection of the ascending aorta and aortic arch are often associated conditions that complicate patient management and increase morbidity and mortality. These associated aortopathies are commonly attributed to turbulent hemodynamic flow through the malformed valve leading to focal defects in the vessel wall. However, numerous surgical and pathological studies have identified widespread cystic medial necrosis and smooth muscle apoptosis throughout the aortic arch in affected patients. Here, we provide experimental evidence for an alternative model to explain the association of aortic vessel and valvular disease. Using mice with primary and secondary cardiac neural crest deficiencies, we have shown that neural crest contribution to the outflow endocardial cushions (the precursors of the semilunar valves) is required for late gestation valvular remodeling, mesenchymal apoptosis, and proper valve architecture. Neural crest was also shown to contribute to the smooth muscle layer of the wall of the ascending aorta and aortic arch. Hence, defects of cardiac neural crest can result in functionally abnormal semilunar valves and concomitant aortic arch artery abnormalities.
Figures




References
-
- Nakajima Y, Yamagishi T, Hokari S, Nakamura H. Mechanisms involved in valvuloseptal endocardial cushion formation in early cardiogenesis: roles of transforming growth factor (TGF)-beta and bone morphogenetic protein (BMP). Anat Rec. 2000;258(2):119–127. doi: 10.1002/(SICI)1097-0185(20000201)258:2<119::AID-AR1>3.0.CO;2-U. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
