

HIV type 1 Env precursor cleavage state affects recognition by both neutralizing and nonneutralizing gp41 antibodies
- PMID: 21158699
- PMCID: PMC3159124
- DOI: 10.1089/AID.2010.0281
HIV type 1 Env precursor cleavage state affects recognition by both neutralizing and nonneutralizing gp41 antibodies
Abstract
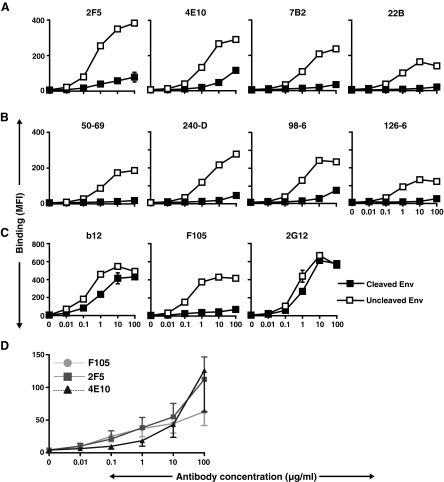
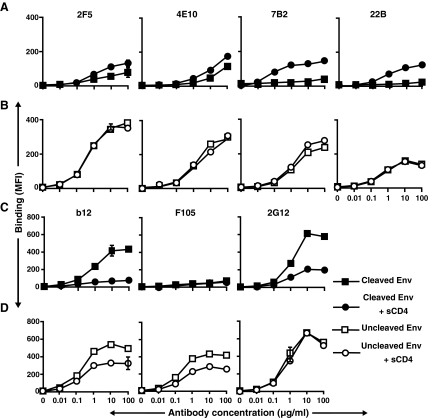
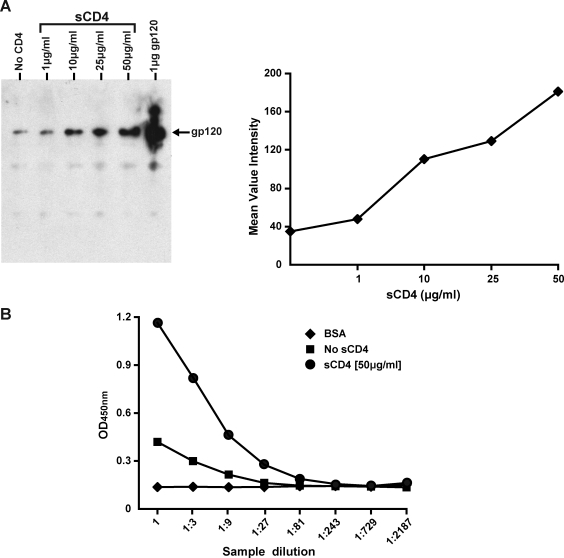
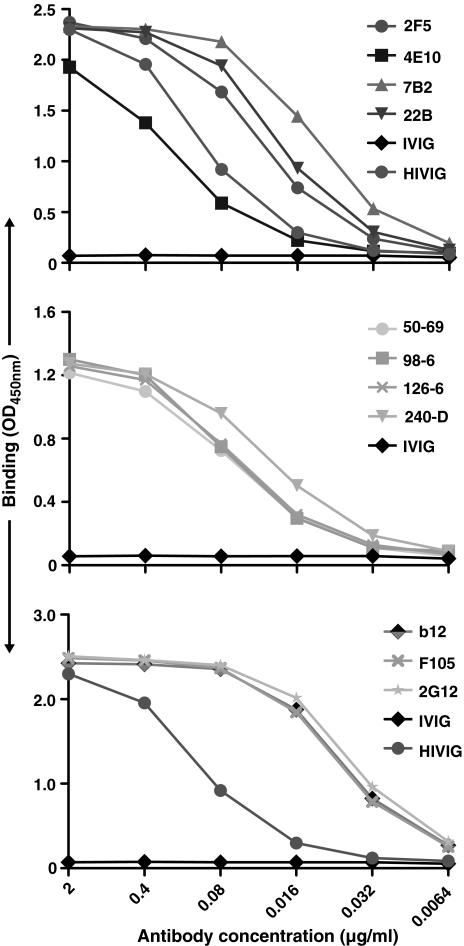
HIV-1 is relatively resistant to antibody-mediated neutralization; however, rare antibodies to the exterior envelope glycoprotein, gp120, and the transmembrane glycoprotein, gp41, can neutralize a broad array of isolates. Two antibodies, 2F5 and 4E10, are directed against the gp41 membrane proximal external region (MPER); however, the kinetic neutralization signature of these antibodies remains unresolved. Previously, we reported that the fully cleaved, cell surface envelope glycoproteins (Env) derived from the primary isolate, JR-FL, are well recognized exclusively by gp120-directed neutralizing ligands and not by nonneutralizing gp120 antibodies. However, the gp120 nonneutralizing antibodies can recognize HIV spikes that are rendered fully cleavage defective by site-directed mutagenesis. Here, we extended such analysis to gp41 neutralizing and nonneutralizing antibodies and, relative to the rules of gp120-specific antibody recognition, we observed marked contrasts. Similar to gp120 recognition, the nonneutralizing gp41 cluster 1 or cluster 2 antibodies bound much more efficiently to cleavage-defective spikes when compared to their recognition of cleaved spikes. In contrast to gp120 neutralizing antibody recognition, the broadly neutralizing gp41 antibodies 2F5 and 4E10, like the nonneutralizing gp41 antibodies, did not efficiently recognize the predominantly cleaved, primary isolate JR-FL spikes. However, if the spikes were rendered cleavage defective, recognition by both the neutralizing and nonneutralizing ligand markedly increased. CD4 interaction with the cleaved spikes markedly increased recognition by most nonneutralizing gp41 antibodies, whereas such treatment had a minimal increase of 2F5 and 4E10 recognition. These data indicate again the profound influence that cleavage imposes on the quaternary packing of primary isolate spikes and have important implications for soluble trimer candidate immunogens.
Figures
References
-
- Choe H. Farzan M. Sun Y, et al. The beta-chemokine receptors CCR3 and CCR5 facilitate infection by primary HIV-1 isolates. Cell. 1996;85(7):1135–1148. - PubMed
-
- Dalgleish AG. Beverley PC. Clapham PR. Crawford DH. Greaves MF. Weiss RA. The CD4 (T4) antigen is an essential component of the receptor for the AIDS retrovirus. Nature. 1984;312(5996):763–767. - PubMed
-
- Hallenberger S. Bosch V. Angliker H. Shaw E. Klenk HD. Garten W. Inhibition of furin-mediated cleavage activation of HIV-1 glycoprotein gp160. Nature. 1992;360(6402):358–361. - PubMed
-
- Weissenhorn W. Dessen A. Harrison SC. Skehel JJ. Wiley DC. Atomic structure of the ectodomain from HIV-1 gp41. Nature. 1997;387(6631):426–430. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
