

Functional analysis of Toll-related genes in Drosophila
- PMID: 21158756
- PMCID: PMC4017902
- DOI: 10.1111/j.1440-169X.2010.01213.x
Functional analysis of Toll-related genes in Drosophila
Abstract
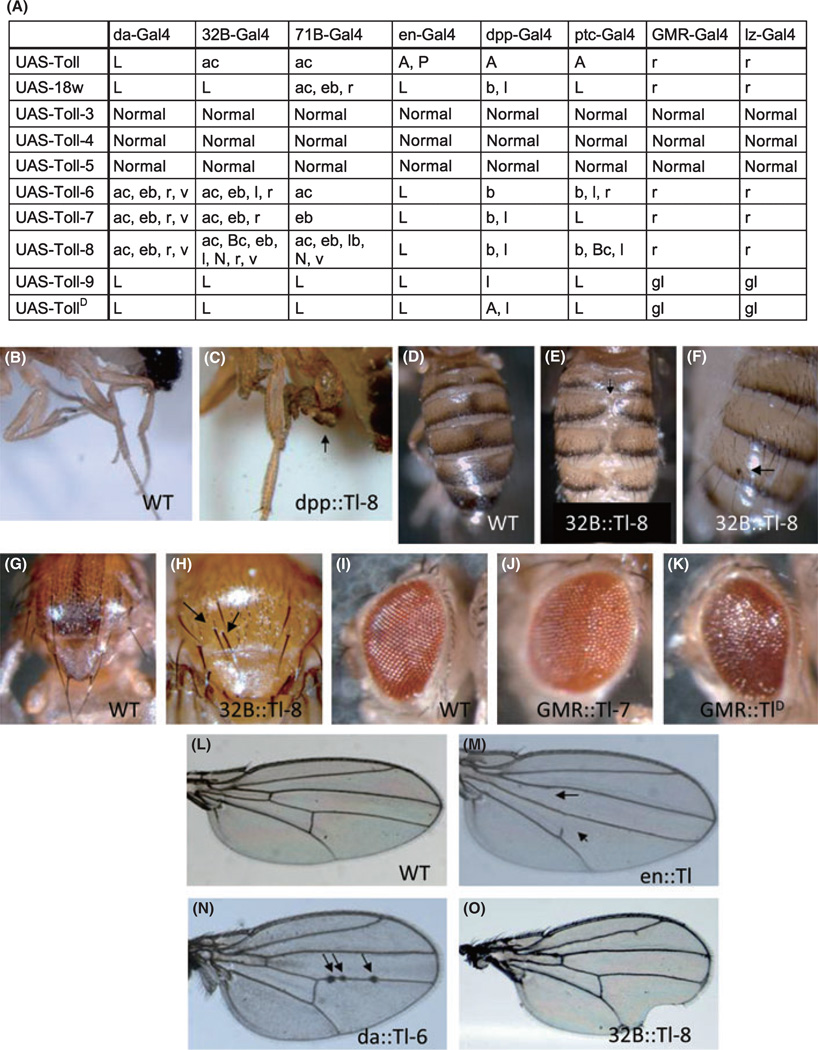
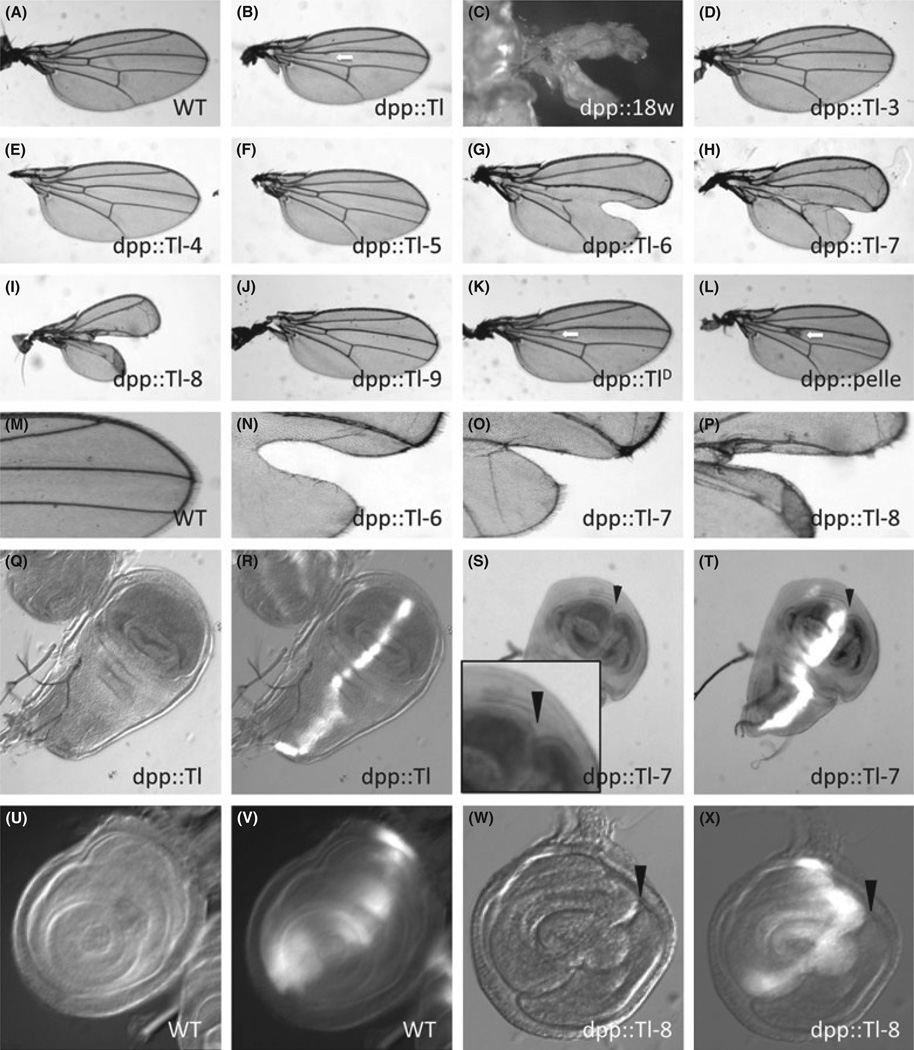
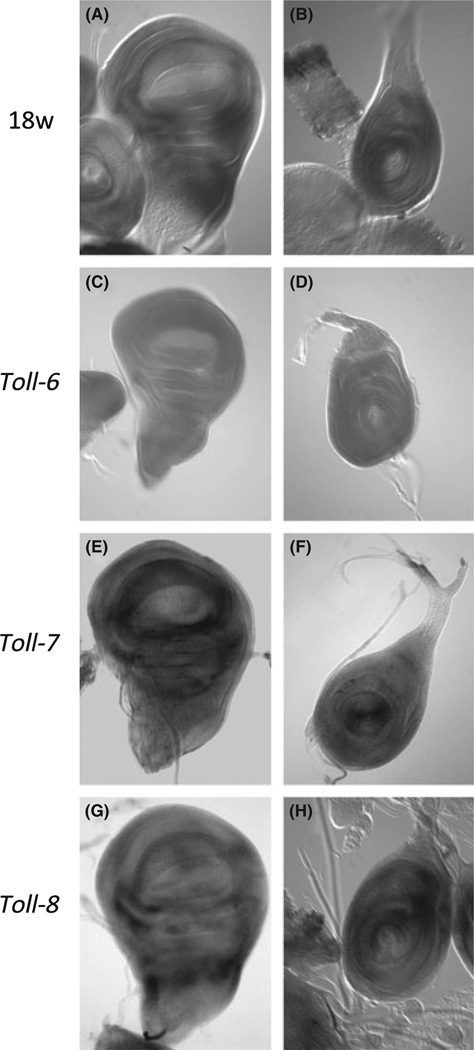
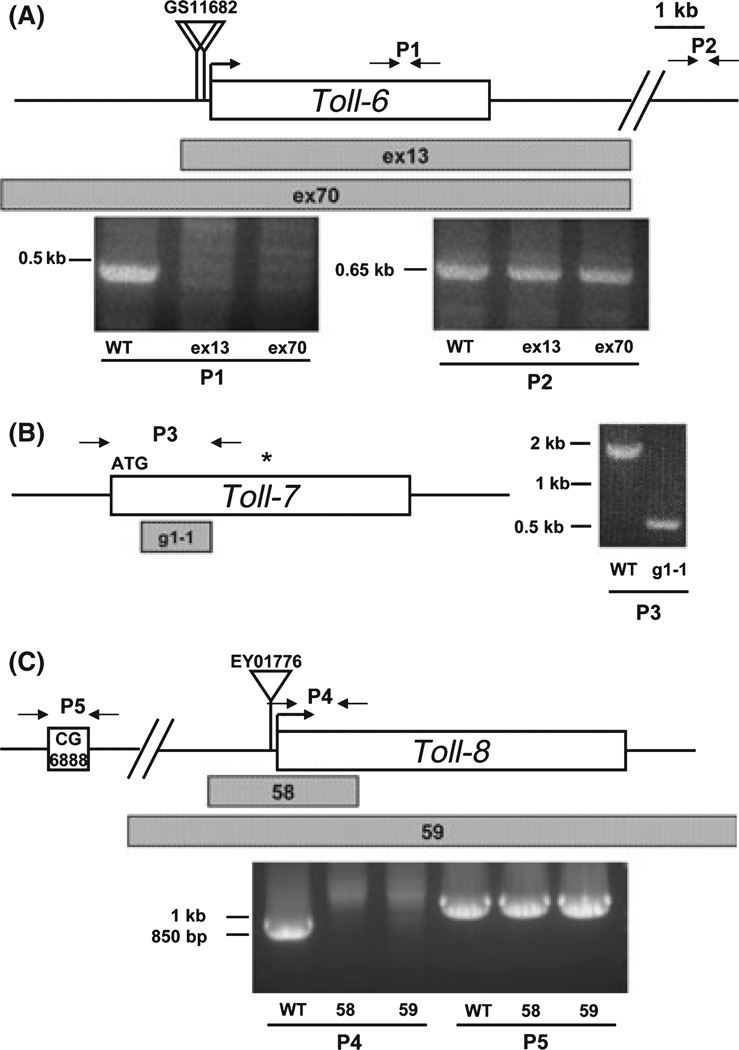
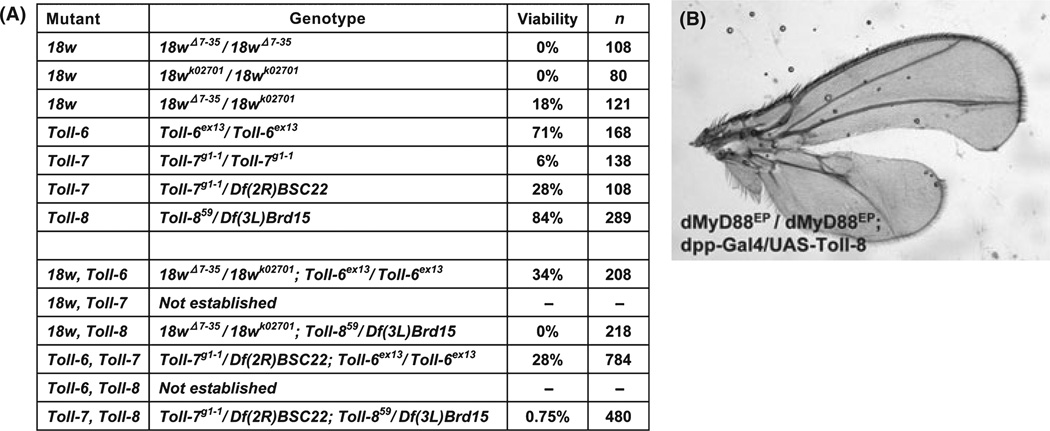
The Drosophila genome encodes a total of nine Toll and related proteins. The immune and developmental functions of Toll and 18Wheeler (18W) have been analyzed extensively, while the in vivo functions of the other Toll-related proteins require further investigation. We performed transgenic experiments and found that overexpression of Toll-related genes caused different extents of lethality and developmental defects. Moreover, 18w, Toll-6, Toll-7 and Toll-8 often caused related phenotypic changes, consistent with the idea that these four genes have more conserved molecular structure and thus may regulate similar processes in vivo. Deletion alleles of Toll-6, Toll-7 and Toll-8 were generated by targeted homologous recombination or P element excision. These mutant alleles were viable, fertile, and had no detectable defect in the inducible expression of antimicrobial peptide genes except for the Toll-8 mutant had some defects in leg development. The expression of 18w, Toll-7 and Toll-8 mRNA showed wide and overlapping patterns in imaginal discs and the 18w, Toll-8 double and Toll-7, Toll-8 double mutants showed substantially increased lethality. Overall our results suggest that some of the Toll-related proteins, such as 18W, Toll-7 and Toll-8, may have redundant functions in regulating developmental processes.
© 2010 The Authors. Journal compilation © 2010 Japanese Society of Developmental Biologists.
Figures
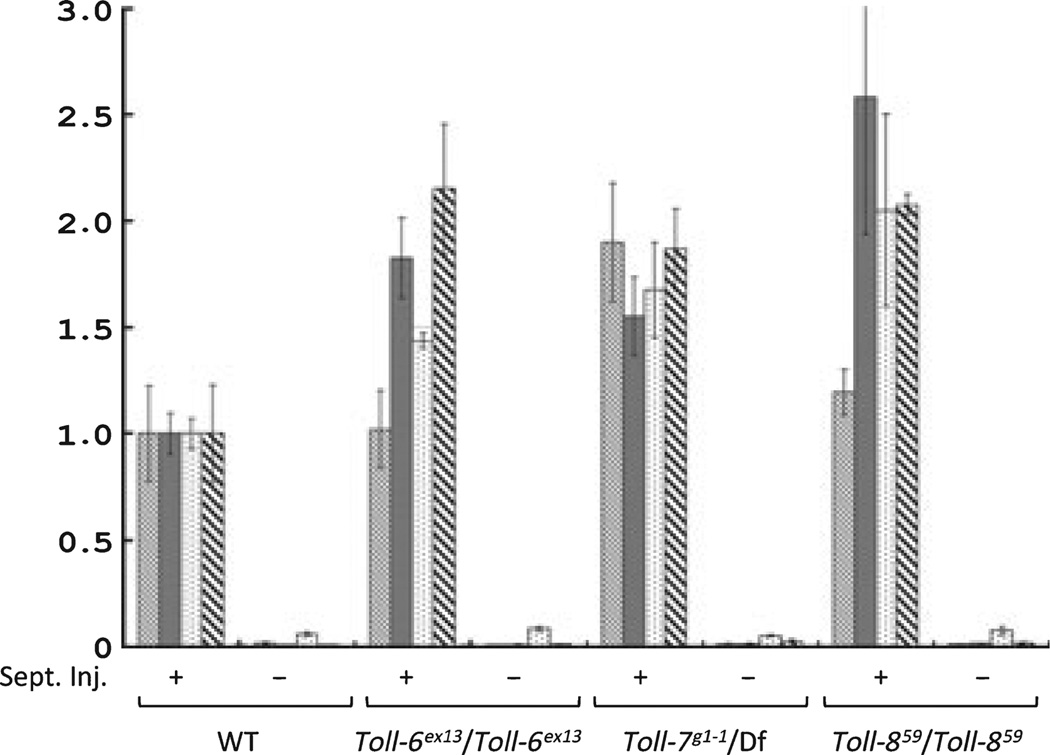
 ); Dipt is Diptericin (
); Dipt is Diptericin ( ); Drs is Drosomycin (
); Drs is Drosomycin ( ); Mtk is Metchnikowin (
); Mtk is Metchnikowin ( ). The inducibility of the antimicrobial peptide genes tested remains high in the mutants as in the wild type flies.
). The inducibility of the antimicrobial peptide genes tested remains high in the mutants as in the wild type flies.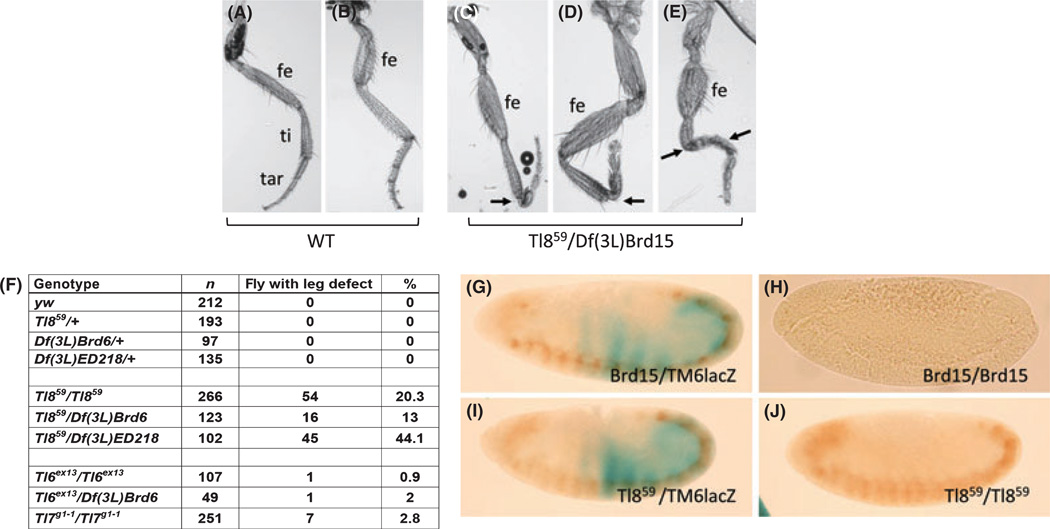
References
-
- Anderson KV, Jurgens G, Nusslein-Volhard C. Establishment of dorsal-ventral polarity in the Drosophila embryo: genetic studies on the role of the Toll gene product. Cell. 1985;42:779–789. - PubMed
-
- Aoki K, Perlman M, Lim JM, Cantu R, Wells L, Tiemeyer M. Dynamic developmental elaboration of N-linked glycan complexity in the Drosophila melanogaster embryo. J. Biol. Chem. 2007;282:9127–9142. - PubMed
-
- Bettencourt R, Tanji T, Yagi Y, Ip YT. Toll and Toll-9 in Drosophila innate immune response. J. Endotoxin Res. 2004;10:261–268. - PubMed
-
- Brennan CA, Anderson KV. Drosophila: the genetics of innate immune recognition and response. Annu. Rev. Immunol. 2004;22:457–483. - PubMed
-
- Cheng TC, Zhang YL, Liu C, Xu PZ, Gao ZH, Xia QY, Xiang ZH. Identification and analysis of Toll-related genes in the domesticated silkworm, Bombyx mori. Dev. Comp. Immunol. 2008;32:464–475. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
