

Encoding of eye position in the goldfish horizontal oculomotor neural integrator
- PMID: 21160010
- PMCID: PMC4103780
- DOI: 10.1152/jn.00313.2010
Encoding of eye position in the goldfish horizontal oculomotor neural integrator
Abstract
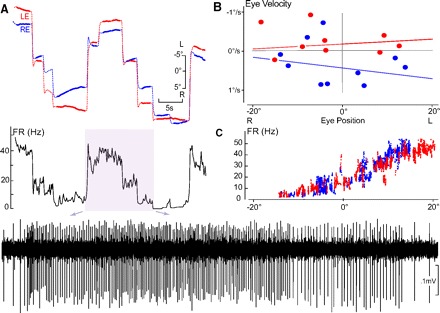
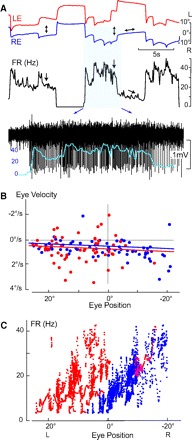
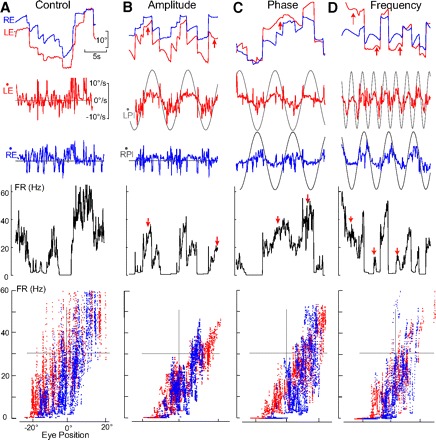
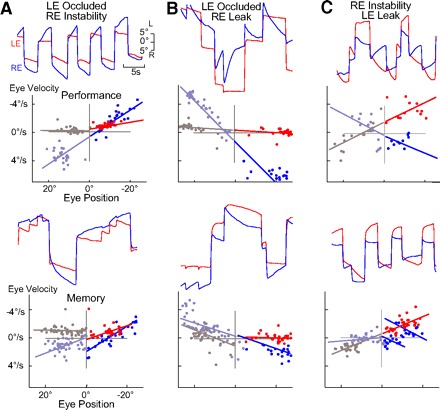
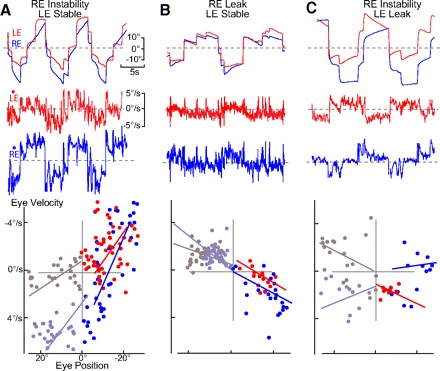
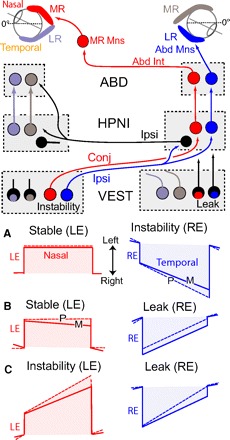
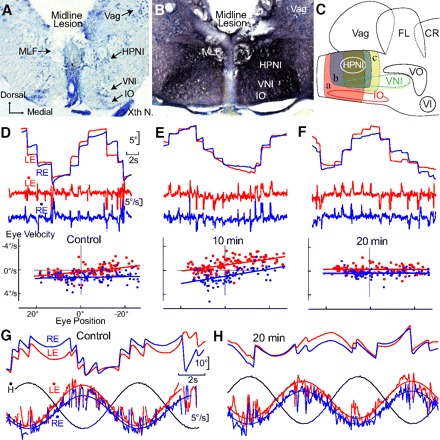
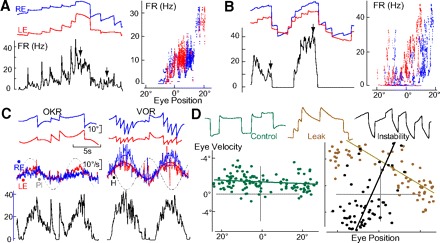
Monocular organization of the goldfish horizontal neural integrator was studied during spontaneous scanning saccadic and fixation behaviors. Analysis of neuronal firing rates revealed a population of ipsilateral (37%), conjugate (59%), and contralateral (4%) eye position neurons. When monocular optokinetic stimuli were employed to maximize disjunctive horizontal eye movements, the sampled population changed to 57, 39, and 4%. Monocular eye tracking could be elicited at different gain and phase with the integrator time constant independently modified for each eye by either centripetal (leak) or centrifugal (instability) drifting visual stimuli. Acute midline separation between the hindbrain oculomotor integrators did not affect either monocularity or time constant tuning, corroborating that left and right eye positions are independently encoded within each integrator. Together these findings suggest that the "ipsilateral" and "conjugate/contralateral" integrator neurons primarily target abducens motoneurons and internuclear neurons, respectively. The commissural pathway is proposed to select the conjugate/contralateral eye position neurons and act as a feedforward inhibition affecting null eye position, oculomotor range, and saccade pattern.
Figures
References
-
- Aksay E, Baker R, Seung HS, Tank DW. Anatomy and discharge properties of pre-motor neurons in the goldfish medulla that have eye-position signals during fixations. J Neurophysiol 84: 1035–1049, 2000. - PubMed
-
- Aksay E, Gamkrelidze G, Seung HS, Baker R, Tank DW. In vivo intracellular recording and perturbation of persistent activity in a neural integrator. Nat Neurosci 4: 184–193, 2001. - PubMed
-
- Aksay E, Major G, Goldman MS, Baker R, Seung HS, Tank DW. History dependence of rate covariation between neurons during persistent activity in an oculomotor integrator. Cereb Cortex 13: 1173–1184, 2003. - PubMed
-
- Allum JH, Graf W, Dichgans J, Schmidt CL. Visual-vestibular interactions in the vestibular nuclei of the goldfish. Exp Brain Res 26: 463–485, 1976. - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources