

Disruption of Adult Neurogenesis in the Olfactory Bulb Affects Social Interaction but not Maternal Behavior
- PMID: 21160552
- PMCID: PMC3001759
- DOI: 10.3389/fnbeh.2010.00176
Disruption of Adult Neurogenesis in the Olfactory Bulb Affects Social Interaction but not Maternal Behavior
Abstract
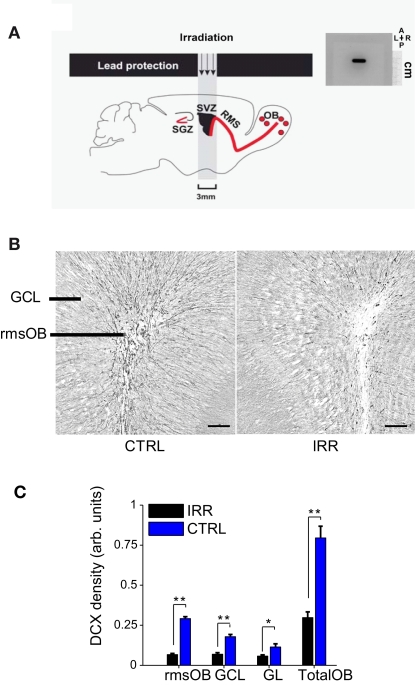
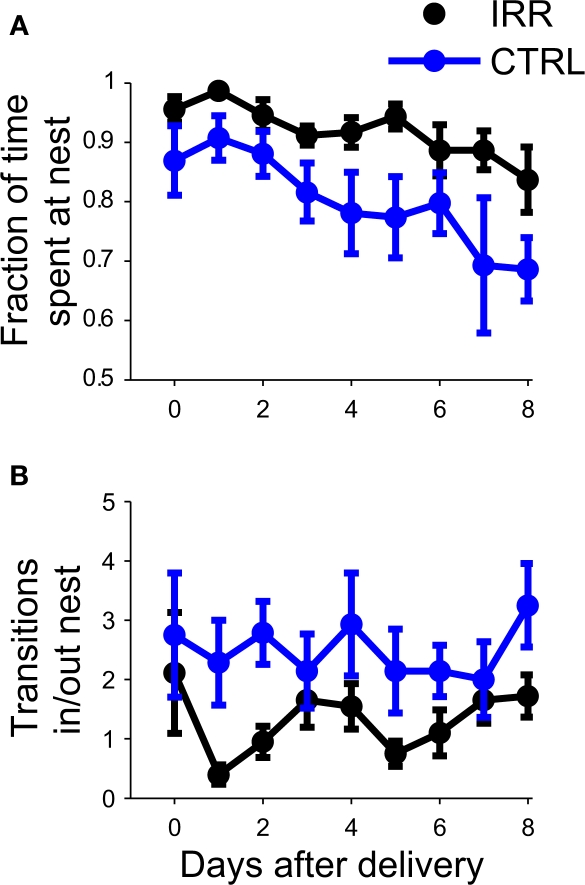
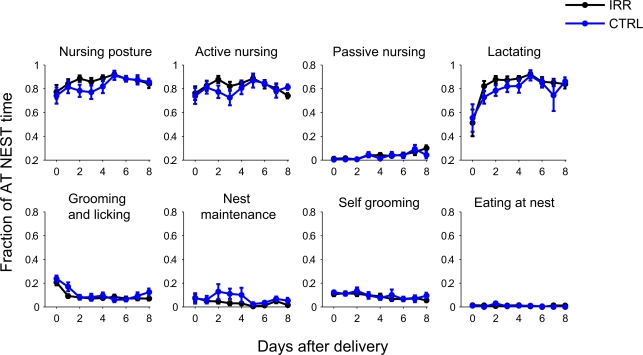
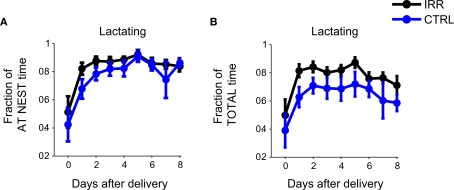
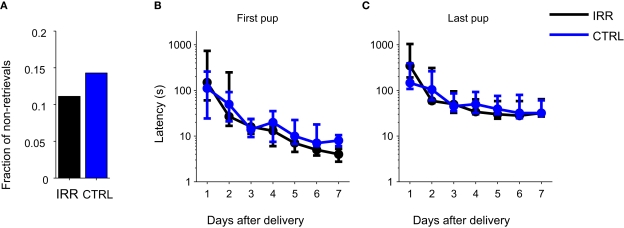
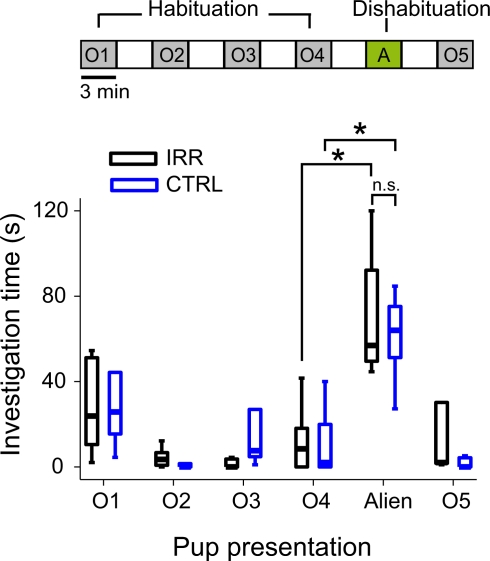
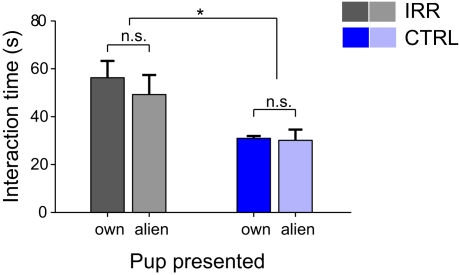
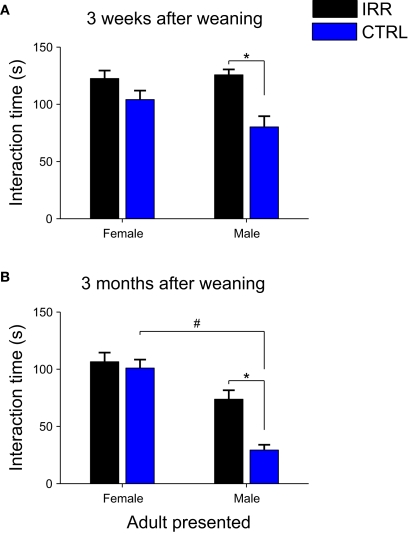
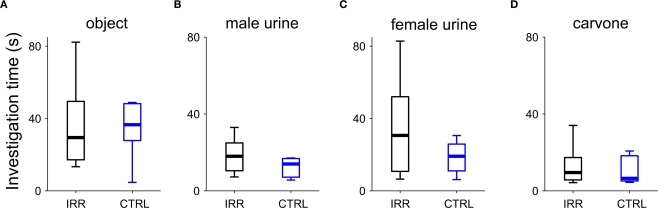
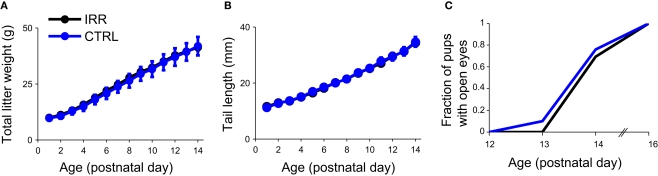
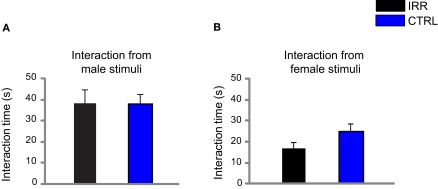
Adult-born neurons arrive to the olfactory bulb (OB) and integrate into the existing circuit throughout life. Despite the prevalence of this phenomenon, its functional impact is still poorly understood. Recent studies point to the importance of newly generated neurons to olfactory learning and memory. Adult neurogenesis is regulated by a variety of factors, notably by instances related to reproductive behavior, such as exposure to mating partners, pregnancy and lactation, and exposure to offspring. To study the contribution of olfactory neurogenesis to maternal behavior and social recognition, here we selectively disrupted OB neurogenesis using focal irradiation of the subventricular zone in adult female mice. We show that reduction of olfactory neurogenesis results in an abnormal social interaction pattern with male, but not female, conspecifics; we suggest that this effect could result from the inability to detect or discriminate male odors and could therefore have implications for the recognition of potential mating partners. Disruption of OB neurogenesis, however, neither impaired maternal-related behaviors, nor did it affect the ability of mothers to discriminate their own progeny from others.
Keywords: irradiation; mate recognition; maternal behavior; neurogenesis; offspring recognition; olfaction; social interaction.
Figures
References
-
- Abraham N. M., Egger V., Shimshek D. R., Renden R., Fukunaga I., Sprengel R., Seeburg P. H., Klugmann M., Margrie T. W., Schaefer A. T., Kuner T. (2010). Synaptic inhibition in the olfactory bulb accelerates odor discrimination in mice. Neuron 65, 399–41110.1016/j.neuron.2010.01.009 - DOI - PMC - PubMed
-
- Barnard C. J., Fitzsimons J. (1988). Kin recognition and mate choice in mice: the effects of kinship, familiarity and social interference on intersexual interaction. Anim. Behav. 36, 1078–109010.1016/S0003-3472(88)80067-8 - DOI
LinkOut - more resources
Full Text Sources
Miscellaneous