

Endotoxin activates de novo sphingolipid biosynthesis via nuclear factor kappa B-mediated upregulation of Sptlc2
- PMID: 21167294
- PMCID: PMC3366150
- DOI: 10.1016/j.prostaglandins.2010.12.003
Endotoxin activates de novo sphingolipid biosynthesis via nuclear factor kappa B-mediated upregulation of Sptlc2
Abstract
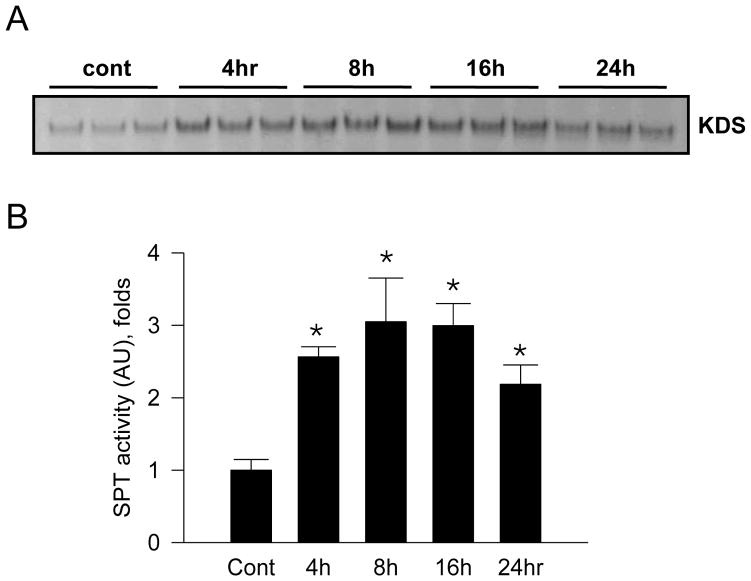
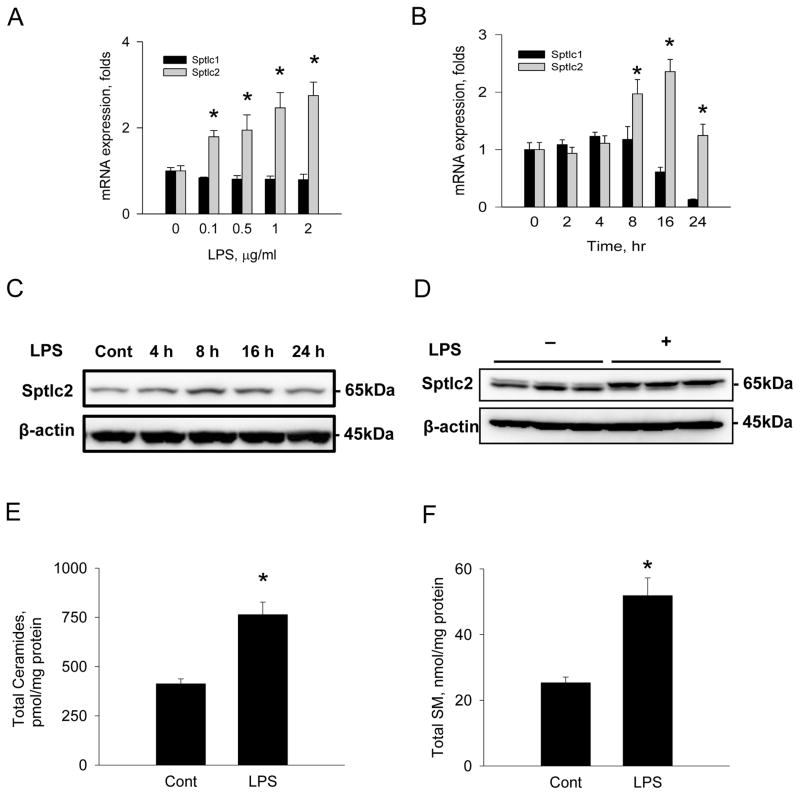
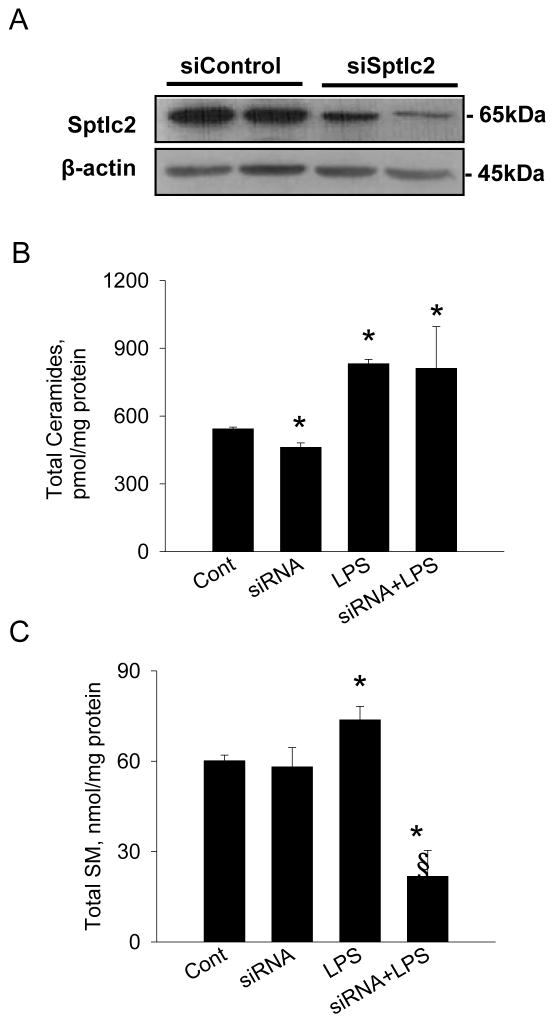
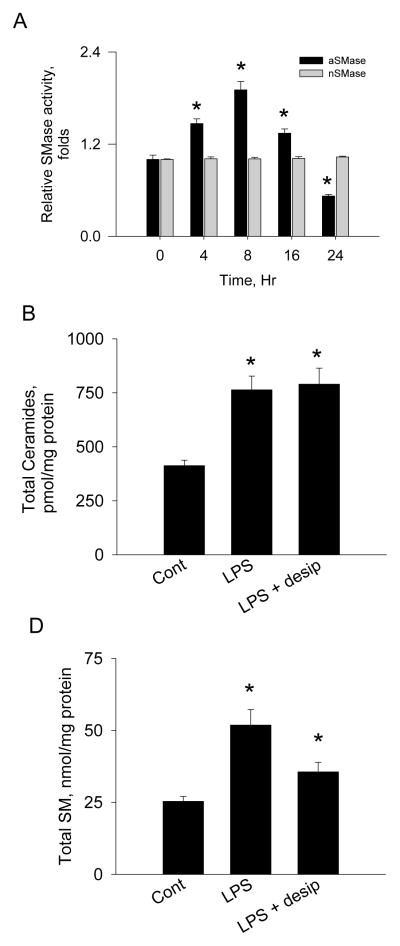
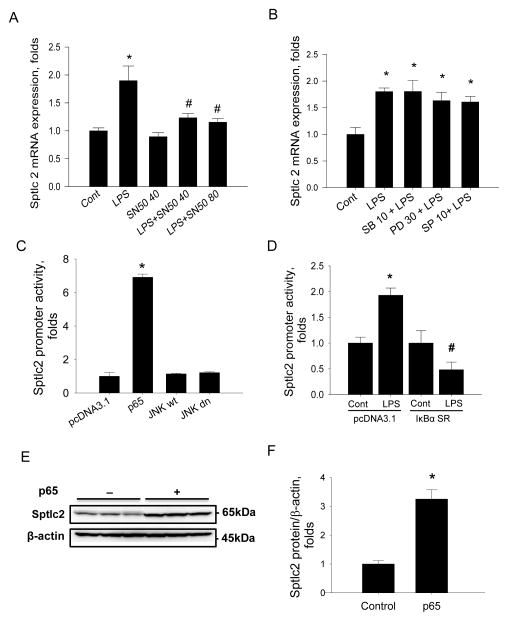
Sphingolipids are membrane components and are involved in cell proliferation, apoptosis and metabolic regulation. In this study we investigated whether de novo sphingolipid biosynthesis in macrophages is regulated by inflammatory stimuli. Lipopolysaccharide (LPS) treatment upregulated Sptlc2, a subunit of serine palmitoyltransferase (SPT), mRNA and protein in Raw264.7 and mouse peritoneal macrophages, but Sptlc1, another subunit of SPT, was not altered. SPT activation by LPS elevated cellular levels of ceramides and sphingomyelin (SM). Pharmacological inhibition of nuclear factor kappa B (NFκB) prevented LPS-induced upregulation of Sptlc2 while transfection of p65 subunit of NFκB upregulated Sptlc2 and increased cellular ceramide levels. In contrast, MAP kinases were not involved in regulation of sphingolipid biosynthesis. Analysis of Sptlc2 promoter and chromatin immunoprecipitation (ChIP) assay showed that NFκB binding sites are located in Sptlc2 promoter region. Our results demonstrate that inflammatory stimuli activate de novo sphingolipid biosynthesis via NFκB and may play a critical role in lipid metabolism in macrophages.
Crown Copyright © 2010. Published by Elsevier Inc. All rights reserved.
Figures

References
-
- Pettus BJ, Chalfant CE, Hannun YA. Ceramide in apoptosis: an overview and current perspectives. Biochim Biophys Acta. 2002;1585:114–25. - PubMed
-
- Chavez JA, Summers SA. Characterizing the effects of saturated fatty acids on insulin signaling and ceramide and diacylglycerol accumulation in 3T3-L1 adipocytes and C2C12 myotubes. Arch Biochem Biophys. 2003;419:101–9. - PubMed
-
- Park TS, Panek RL, Mueller SB, Hanselman JC, Rosebury WS, Robertson AW, et al. Inhibition of sphingomyelin synthesis reduces atherogenesis in apolipoprotein E-knockout mice. Circulation. 2004;110:3465–71. - PubMed
-
- Holland WL, Brozinick JT, Wang LP, Hawkins ED, Sargent KM, Liu Y, et al. Inhibition of ceramide synthesis ameliorates glucocorticoid-, saturated-fat-, and obesity-induced insulin resistance. Cell Metab. 2007;5:167–79. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
