

The dorsal medial frontal cortex is sensitive to time on task, not response conflict or error likelihood
- PMID: 21168515
- PMCID: PMC3114292
- DOI: 10.1016/j.neuroimage.2010.12.027
The dorsal medial frontal cortex is sensitive to time on task, not response conflict or error likelihood
Abstract
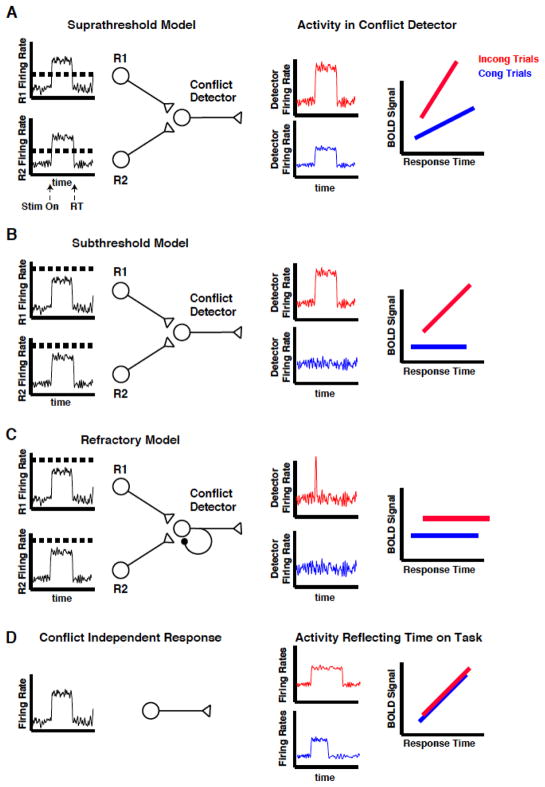
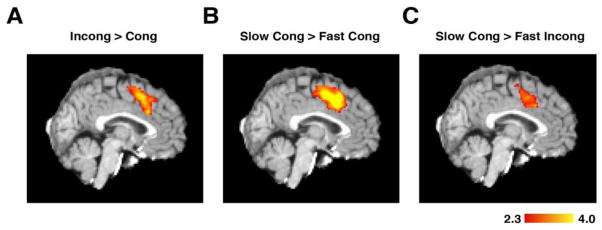
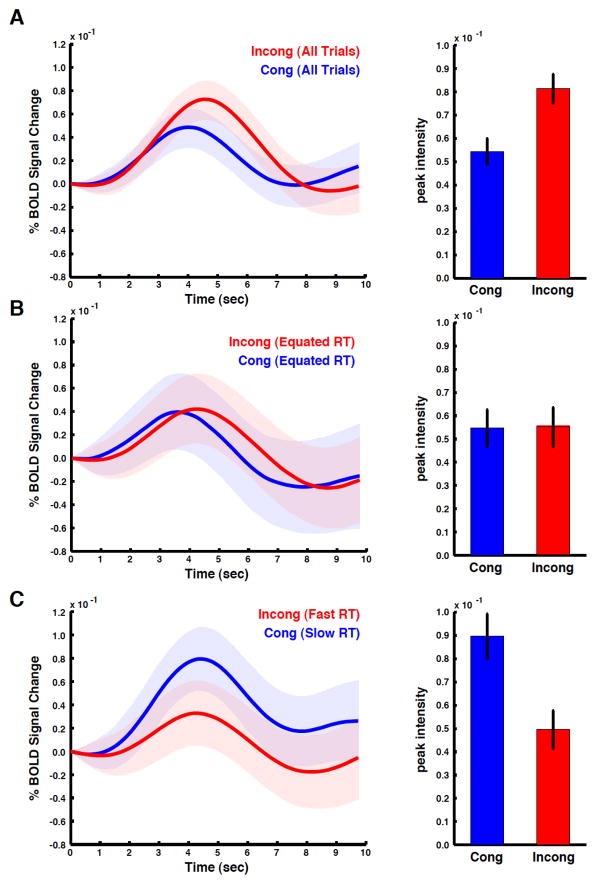
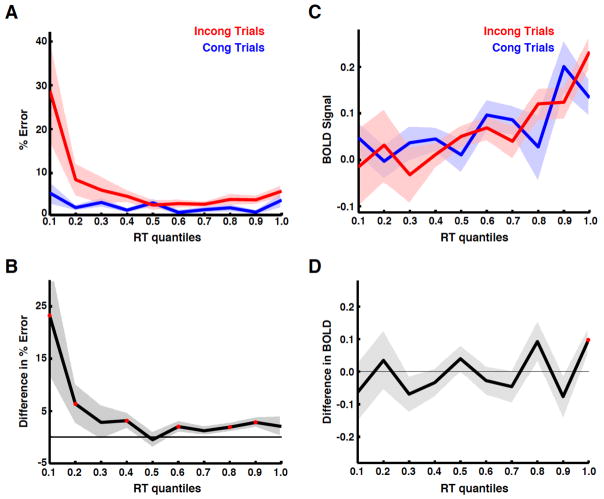
The dorsal medial frontal cortex (dMFC) is highly active during choice behavior. Though many models have been proposed to explain dMFC function, the conflict monitoring model is the most influential. It posits that dMFC is primarily involved in detecting interference between competing responses thus signaling the need for control. It accurately predicts increased neural activity and response time (RT) for incompatible (high-interference) vs. compatible (low-interference) decisions. However, it has been shown that neural activity can increase with time on task, even when no decisions are made. Thus, the greater dMFC activity on incompatible trials may stem from longer RTs rather than response conflict. This study shows that (1) the conflict monitoring model fails to predict the relationship between error likelihood and RT, and (2) the dMFC activity is not sensitive to congruency, error likelihood, or response conflict, but is monotonically related to time on task.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
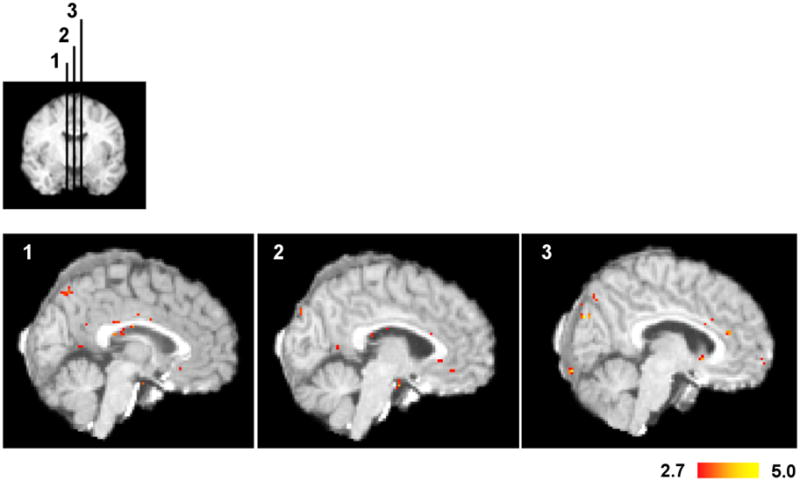
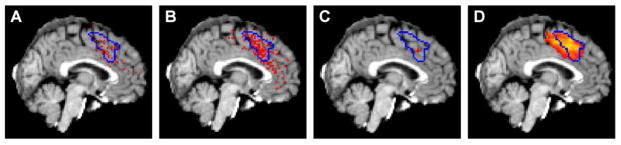
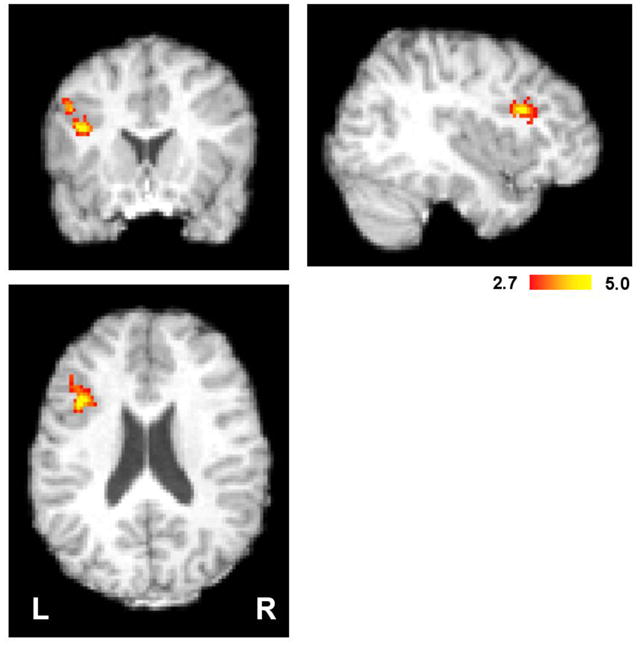
Comment in
-
Errors of interpretation and modeling: a reply to Grinband et al.Neuroimage. 2011 Jul 15;57(2):316-9. doi: 10.1016/j.neuroimage.2011.04.029. Epub 2011 Apr 22. Neuroimage. 2011. PMID: 21530662 Free PMC article.
-
The blind executive.Neuroimage. 2011 Jul 15;57(2):312-3. doi: 10.1016/j.neuroimage.2011.04.025. Epub 2011 Apr 22. Neuroimage. 2011. PMID: 21540115 No abstract available.
-
Medial prefrontal cortex activity correlates with time-on-task: what does this tell us about theories of cognitive control?Neuroimage. 2011 Jul 15;57(2):314-5. doi: 10.1016/j.neuroimage.2011.04.028. Epub 2011 Apr 22. Neuroimage. 2011. PMID: 21540116
References
-
- Banich MT, Milham MP, Atchley R, Cohen NJ, Webb A, Wszalek T, Kramer AF, Liang ZP, Wright A, Shenker J, Magin R. fMri studies of Stroop tasks reveal unique roles of anterior and posterior brain systems in attentional selection. J Cogn Neurosci. 2000;12:988–1000. - PubMed
-
- Botvinick M, Nystrom LE, Fissell K, Carter CS, Cohen JD. Conflict monitoring versus selection-for-action in anterior cingulate cortex. Nature. 1999;402:179–181. - PubMed
-
- Botvinick MM, Braver TS, Barch DM, Carter CS, Cohen JD. Conflict monitoring and cognitive control. Psychol Rev. 2001;108:624–652. - PubMed
-
- Brown JW, Braver TS. Learned predictions of error likelihood in the anterior cingulate cortex. Science. 2005;307:1118–1121. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
