

doi: 10.1016/j.chembiol.2010.09.015.
Photoreactive stapled BH3 peptides to dissect the BCL-2 family interactome
Affiliations
- PMID: 21168768
- PMCID: PMC3048092
- DOI: 10.1016/j.chembiol.2010.09.015
Item in Clipboard
Photoreactive stapled BH3 peptides to dissect the BCL-2 family interactome
Chem Biol.
.
Abstract
Defining protein interactions forms the basis for discovery of biological pathways, disease mechanisms, and opportunities for therapeutic intervention. To harness the robust binding affinity and selectivity of structured peptides for interactome discovery, we engineered photoreactive stapled BH3 peptide helices that covalently capture their physiologic BCL-2 family targets. The crosslinking α helices covalently trap both static and dynamic protein interactors, and enable rapid identification of interaction sites, providing a critical link between interactome discovery and targeted drug design.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures

(A) Photoreactive stabilized alpha-helix of BCL-2 domains (pSAHBs) for protein capture are generated by replacing native aromatic residues with 4-benzoylphenylalanine (Bpa) followed by ring closing metathesis of olefinic non-natural amino acids installed at i, i +4 positions. (B) A panel of pSAHBs was modeled after the BH3 death domain of pro-apoptotic BAD. (C) BAD pSAHBs retain high affinity binding to anti-apoptotic BCL-XLΔC, as measured by fluorescence polarization assay. Error bars represent mean +/− s.d. for binding assays performed in at least triplicate. (D) Select fluorescently labeled BAD pSAHBs crosslink to BCL-XLΔC upon exposure to UV irradiation, as detected by coomassie stain and fluorescence imaging. (E) Time-course for UV-dependent crosslinking of BAD pSAHB-2 to BCL-XLΔC as monitored by fluorescence scan and coomassie stain, and the corresponding densitometry-based calculation of crosslinking yield using ImageJ software.

(A) The selectivity of BAD pSAHB covalent capture is highlighted by the absence of crosslinking to bovine serum albumin (BSA) and the inability of BSA to disrupt FITC-BAD pSAHB-2/BCL-XLΔC crosslinking. (B) Biotinylated BAD pSAHB-2 was incubated with an equimolar mixture of homologous anti-apoptotic proteins at the indicated ratios in the presence of UV light for 1.5 hours. Western analysis of the isolated crosslinked proteins revealed high fidelity capture of physiologic interactors BCL-XLΔC, BCL-2ΔC, and BCL-wΔC, but not BFL1/A1ΔC, which lies outside the in vivo binding spectrum of the BAD protein. SA, streptavidin. (C) A panel of pSAHBs was modeled after the BIM BH3 domain for comparative crosslinking studies with BAD pSAHBs. (D) BAD and BIM pSAHBs covalently trap their distinct profile of anti-apoptotic binding partners within U937 cellular extracts. (E) In addition to capturing stable protein interactors, BIM pSAHBs 2–4 crosslink to pro-apoptotic BAX, a fleeting “hit and run” target of the BH3-only protein BIM. See also Supplementary Figure 1.
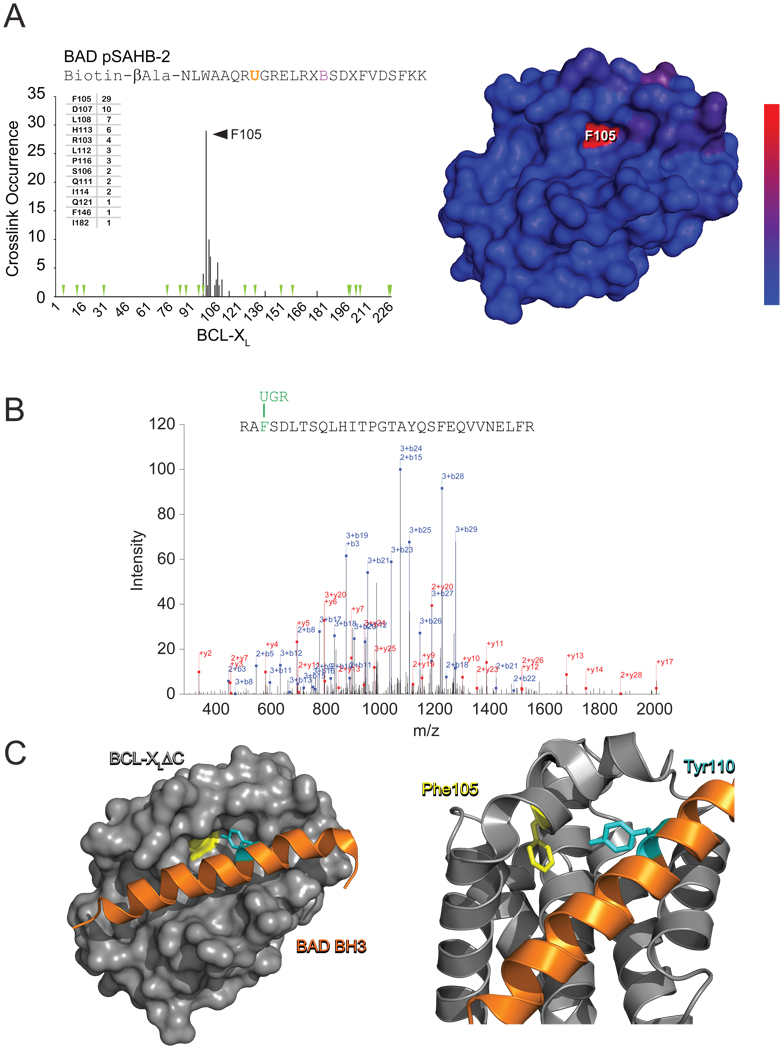
(A) BAD pSAHB-2 was incubated with BCL-XLΔC and the mixture subjected to UV irradiation (1.5 h), electrophoresis, excision of the crosslinked protein, trypsin proteolysis, and LC-MS/MS analysis. The plot depicts the frequency of crosslinked sites identified across the BCL-XLΔC polypeptide sequence. Mapping of the crosslinked residues onto the BCL-XLΔC structure revealed their striking colocalization within a circumscribed region of the canonical BH3-binding pocket, with the frequency of occurrence reflected by the color scale. Green arrowheads, trypsin digestion sites. (B) The predominant Bpa-crosslinked tryptic fragment contained a covalent modification of BCL-XLΔC residue F105, as exemplified by the MS/MS spectrum derived from collision-induced dissociation of the (M+4H)4+ precursor (0.37 parts per million mass accuracy). (C) The most abundant crosslink, located between BAD pSAHB-2 Bpa110 and BCL-XLΔC F105, precisely matches the structurally-defined interaction between BAD BH3 Y110 and BCL-XLΔC F105 at the BH3-binding pocket of BCL-XLΔC (PDB ID 2BZW).

(A, B) BAD pSAHB-2 (A) or BAD pSAHB-4 (B) was incubated with full-length BCL-w and the mixture subjected to UV irradiation, electrophoresis, excision of the crosslinked protein, trypsin proteolysis, and LC-MS/MS analysis. The plots (left) and color scale (right) depict the frequency of BCL-w crosslinked sites, which map to distinct, circumscribed regions within the canonical BH3-binding pocket of BCL-w (right). (C) The top scoring hit for each pSAHB was employed in computational docking analysis to calculate a model structure for the BAD BH3/BCL-w interaction. The BAD BH3 helix engages the canonical BH3-binding pocket of BCL-w with the N to C terminus of the peptide disposed right to left. The en face view (top) demonstrates the complementary electrostatic interactions of D119 and R115 of BAD BH3 with R94 and E84 of BCL-w, respectively. A top view of the structure (bottom) highlights the juxtaposition of crosslinked positions Y110 and F125 (cyan) of the BAD BH3 helix with L63 and R55 of BCL-w, respectively, along the core hydrophobic interface. The side chains of hydrophobic, positively charged, negatively charged, and hydrophilic residues are colored yellow, blue, red and green, respectively. To display an unobstructed view of the BH3-binding site, the BCL-w C-terminus is hidden in the images. (D) The interaction topography of the calculated model structure of BAD BH3/BCL-w is notably similar to that of the structurally-defined BAD BH3/BCL-XLΔC complex (PDB ID 2BZW). The en face view (top) demonstrates the homologous BAD BH3/BCL-XLΔC electrostatic interaction pairs D119/R139 and R115/E129, whereas the core BAD BH3 residues A106, Y110, L114, M117, F121, and F125 of the hydrophobic interface (bottom) are remarkably similar in disposition to these same residues in the calculated model structure of BAD BH3/BCL-w. See also Supplementary Figure 2.
References
-
- Arnesano F, Banci L, Bertini I, Bonvin AM. A docking approach to the study of copper trafficking proteins; interaction between metallochaperones and soluble domains of copper ATPases. Structure. 2004;12:669–676. - PubMed
-
- Blackwell HE, Sadowsky JD, Howard RJ, Sampson JN, Chao JA, Steinmetz WE, O'Leary DJ, Grubbs RH. Ring-closing metathesis of olefinic peptides: design, synthesis, and structural characterization of macrocyclic helical peptides. J Org Chem. 2001;66:5291–5302. - PubMed
-
- Blum JH, Stevens TL, DeFranco AL. Role of the mu immunoglobulin heavy chain transmembrane and cytoplasmic domains in B cell antigen receptor expression and signal transduction. J Biol Chem. 1993;268:27236–27245. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
