

c-Jun NH2-terminal kinase-dependent upregulation of DR5 mediates cooperative induction of apoptosis by perifosine and TRAIL
- PMID: 21172010
- PMCID: PMC3018404
- DOI: 10.1186/1476-4598-9-315
c-Jun NH2-terminal kinase-dependent upregulation of DR5 mediates cooperative induction of apoptosis by perifosine and TRAIL
Abstract
Background: Perifosine, an alkylphospholipid tested in phase II clinical trials, modulates the extrinsic apoptotic pathway and cooperates with tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) to augment apoptosis. The current study focuses on revealing the mechanisms by which perifosine enhances TRAIL-induced apoptosis.
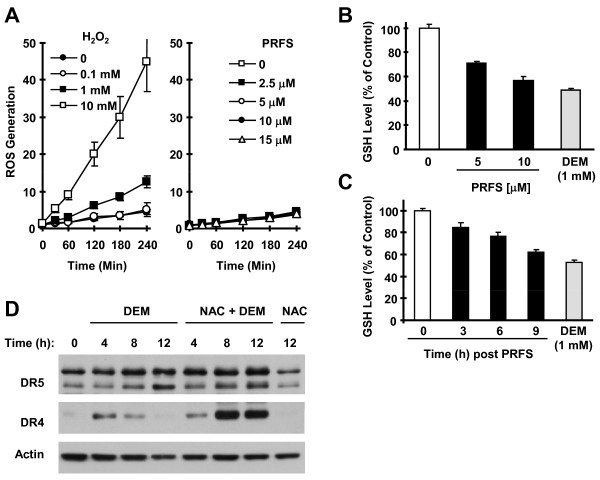
Results: The combination of perifosine and TRAIL was more active than each single agent alone in inducing apoptosis of head and neck squamous cell carcinoma cells and inhibiting the growth of xenografts. Interestingly, perifosine primarily increased cell surface levels of DR5 although it elevated the expression of both DR4 and DR5. Blockade of DR5, but not DR4 upregulation, via small interfering RNA (siRNA) inhibited perifosine/TRAIL-induced apoptosis. Perifosine increased phosphorylated c-Jun NH2-terminal kinase (JNK) and c-Jun levels, which were paralleled with DR4 and DR5 induction. However, only DR5 upregulaiton induced by perifosine could be abrogated by both the JNK inhibitor SP600125 and JNK siRNA. The antioxidants, N-acetylcysteine and glutathione, but not vitamin C or tiron, inhibited perifosine-induced elevation of p-c-Jun, DR4 and DR5. Moreover, no increased production of reactive oxygen species was detected in perifosine-treated cells although reduced levels of intracellular GSH were measured.
Conclusions: DR5 induction plays a critical role in mediating perifosine/TRAIL-induced apoptosis. Perifosine induces DR5 expression through a JNK-dependent mechanism independent of reactive oxygen species.
Figures






Similar articles
-
The alkylphospholipid perifosine induces apoptosis of human lung cancer cells requiring inhibition of Akt and activation of the extrinsic apoptotic pathway.Mol Cancer Ther. 2007 Jul;6(7):2029-38. doi: 10.1158/1535-7163.MCT-07-0004. Epub 2007 Jun 29. Mol Cancer Ther. 2007. PMID: 17604333
-
Quercetin enhances apoptotic effect of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) in ovarian cancer cells through reactive oxygen species (ROS) mediated CCAAT enhancer-binding protein homologous protein (CHOP)-death receptor 5 pathway.Cancer Sci. 2014 May;105(5):520-7. doi: 10.1111/cas.12395. Epub 2014 Apr 11. Cancer Sci. 2014. PMID: 24612139 Free PMC article.
-
Short-hairpin RNA-induced suppression of adenine nucleotide translocase-2 in breast cancer cells restores their susceptibility to TRAIL-induced apoptosis by activating JNK and modulating TRAIL receptor expression.Mol Cancer. 2010 Sep 28;9:262. doi: 10.1186/1476-4598-9-262. Mol Cancer. 2010. PMID: 20875141 Free PMC article.
-
Intracellular localization of DR5 and related regulatory pathways as a mechanism of resistance to TRAIL in cancer.Cell Mol Life Sci. 2017 Jan;74(2):245-255. doi: 10.1007/s00018-016-2321-z. Epub 2016 Aug 10. Cell Mol Life Sci. 2017. PMID: 27510421 Free PMC article. Review.
-
TRAIL-Sensitizing Effects of Flavonoids in Cancer.Int J Mol Sci. 2023 Nov 22;24(23):16596. doi: 10.3390/ijms242316596. Int J Mol Sci. 2023. PMID: 38068921 Free PMC article. Review.
Cited by
-
Gossypol induces apoptosis of multiple myeloma cells through the JUN-JNK pathway.Am J Cancer Res. 2020 Mar 1;10(3):870-883. eCollection 2020. Am J Cancer Res. 2020. PMID: 32266096 Free PMC article.
-
Expression of Death Receptor 4 Is Positively Regulated by MEK/ERK/AP-1 Signaling and Suppressed upon MEK Inhibition.J Biol Chem. 2016 Oct 7;291(41):21694-21702. doi: 10.1074/jbc.M116.738302. Epub 2016 Aug 30. J Biol Chem. 2016. PMID: 27576686 Free PMC article.
-
Verticillin A overcomes apoptosis resistance in human colon carcinoma through DNA methylation-dependent upregulation of BNIP3.Cancer Res. 2011 Nov 1;71(21):6807-16. doi: 10.1158/0008-5472.CAN-11-1575. Epub 2011 Sep 12. Cancer Res. 2011. PMID: 21911457 Free PMC article.
-
Inhibition of mTOR complex 1/p70 S6 kinase signaling elevates PD-L1 levels in human cancer cells through enhancing protein stabilization accompanied with enhanced β-TrCP degradation.Oncogene. 2019 Aug;38(35):6270-6282. doi: 10.1038/s41388-019-0877-4. Epub 2019 Jul 17. Oncogene. 2019. PMID: 31316145
-
Downregulation of death receptor 4 is tightly associated with positive response of EGFR mutant lung cancer to EGFR-targeted therapy and improved prognosis.Theranostics. 2021 Feb 15;11(8):3964-3980. doi: 10.7150/thno.54824. eCollection 2021. Theranostics. 2021. PMID: 33664875 Free PMC article.
References
-
- Kondapaka SB, Singh SS, Dasmahapatra GP, Sausville EA, Roy KK. Perifosine, a novel alkylphospholipid, inhibits protein kinase B activation. Mol Cancer Ther. 2003;2:1093–1103. - PubMed
-
- Chiarini F, Del Sole M, Mongiorgi S, Gaboardi GC, Cappellini A, Mantovani I, Follo MY, McCubrey JA, Martelli AM. The novel Akt inhibitor, perifosine, induces caspase-dependent apoptosis and downregulates P-glycoprotein expression in multidrug-resistant human T-acute leukemia cells by a JNK-dependent mechanism. Leukemia. 2008;22:1106–1116. doi: 10.1038/leu.2008.79. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
