

PML isoforms I and II participate in PML-dependent restriction of HSV-1 replication
- PMID: 21172801
- PMCID: PMC3010193
- DOI: 10.1242/jcs.075390
PML isoforms I and II participate in PML-dependent restriction of HSV-1 replication
Abstract
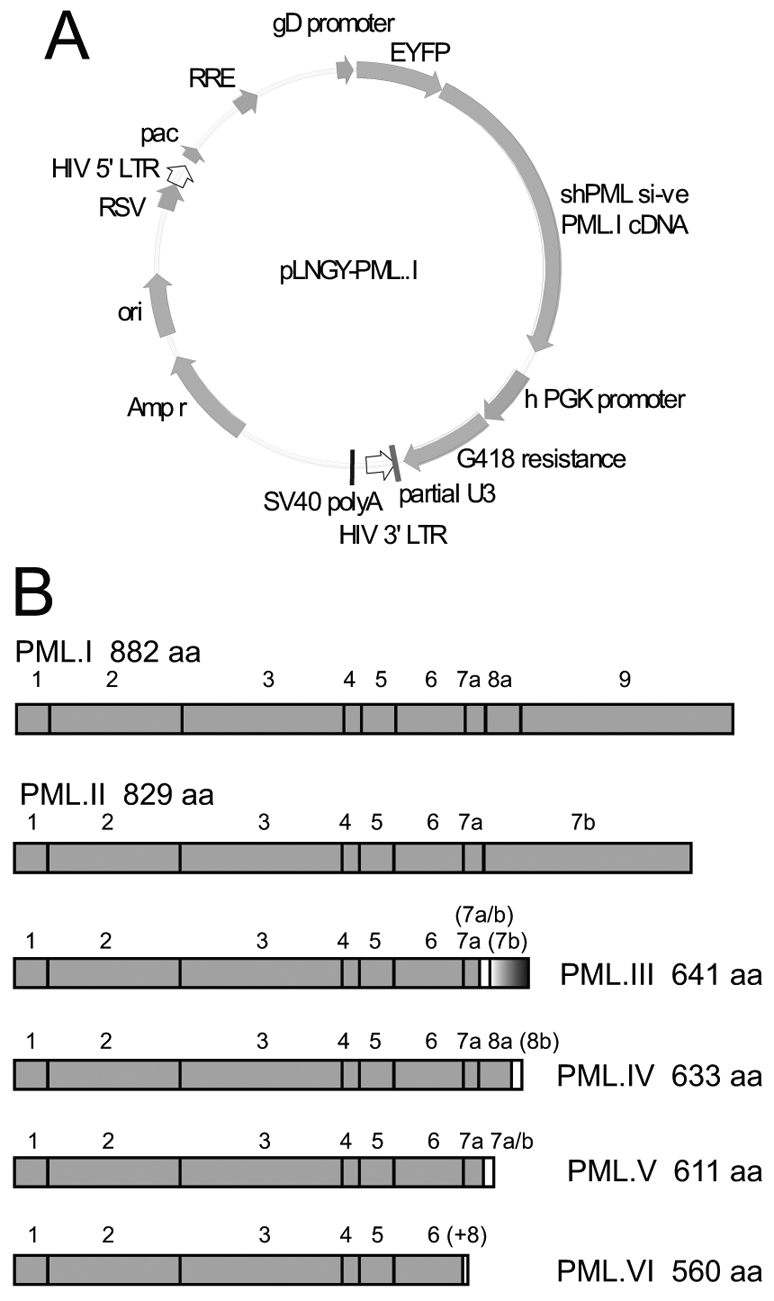
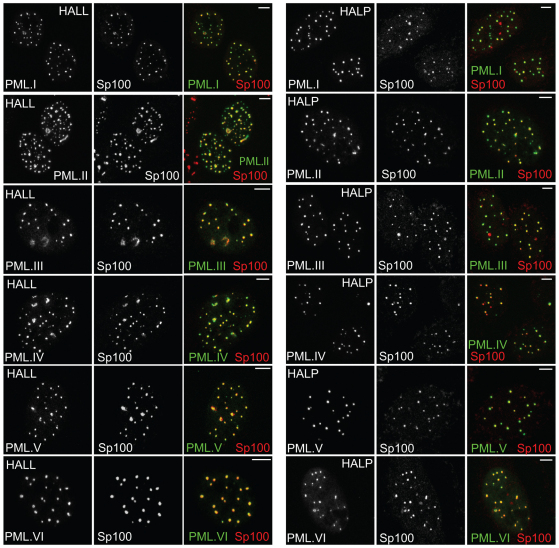
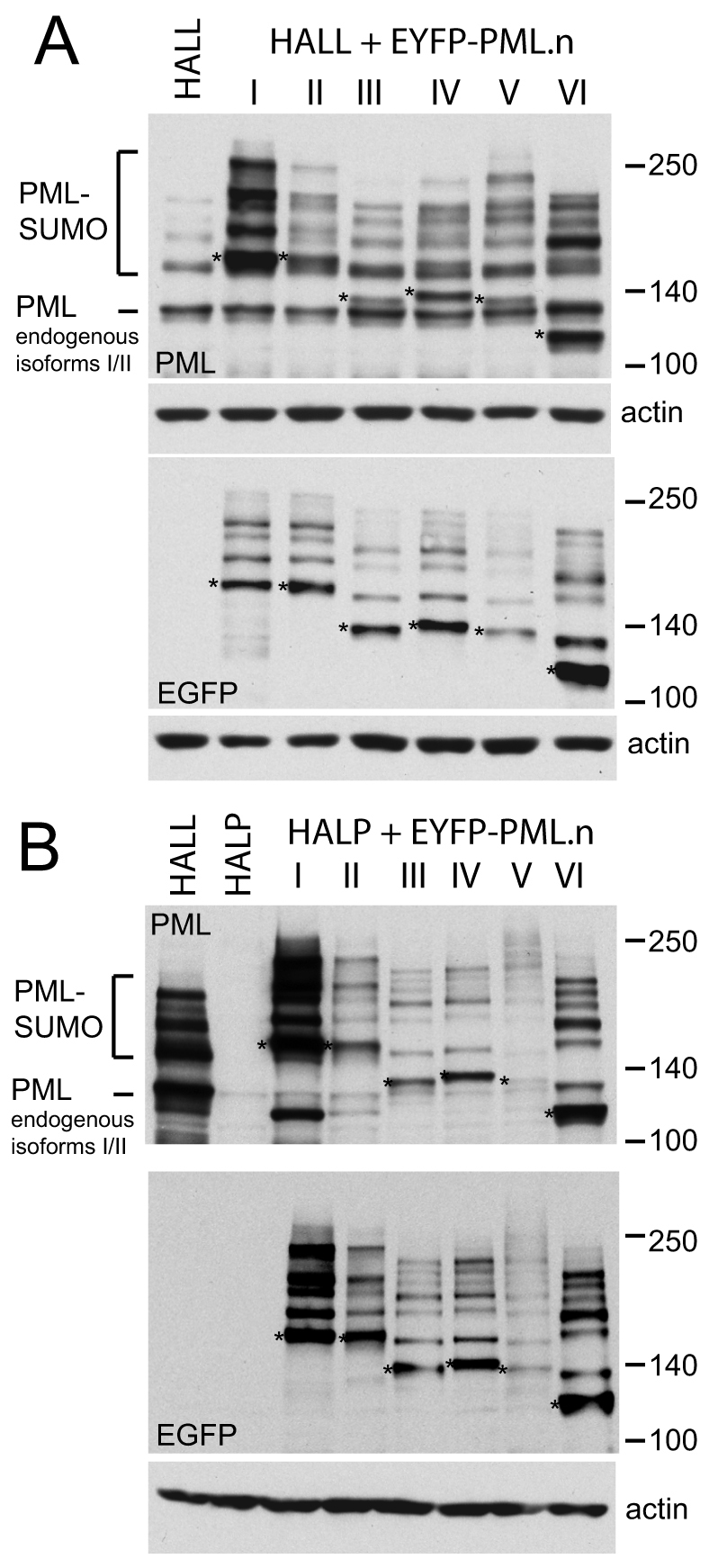
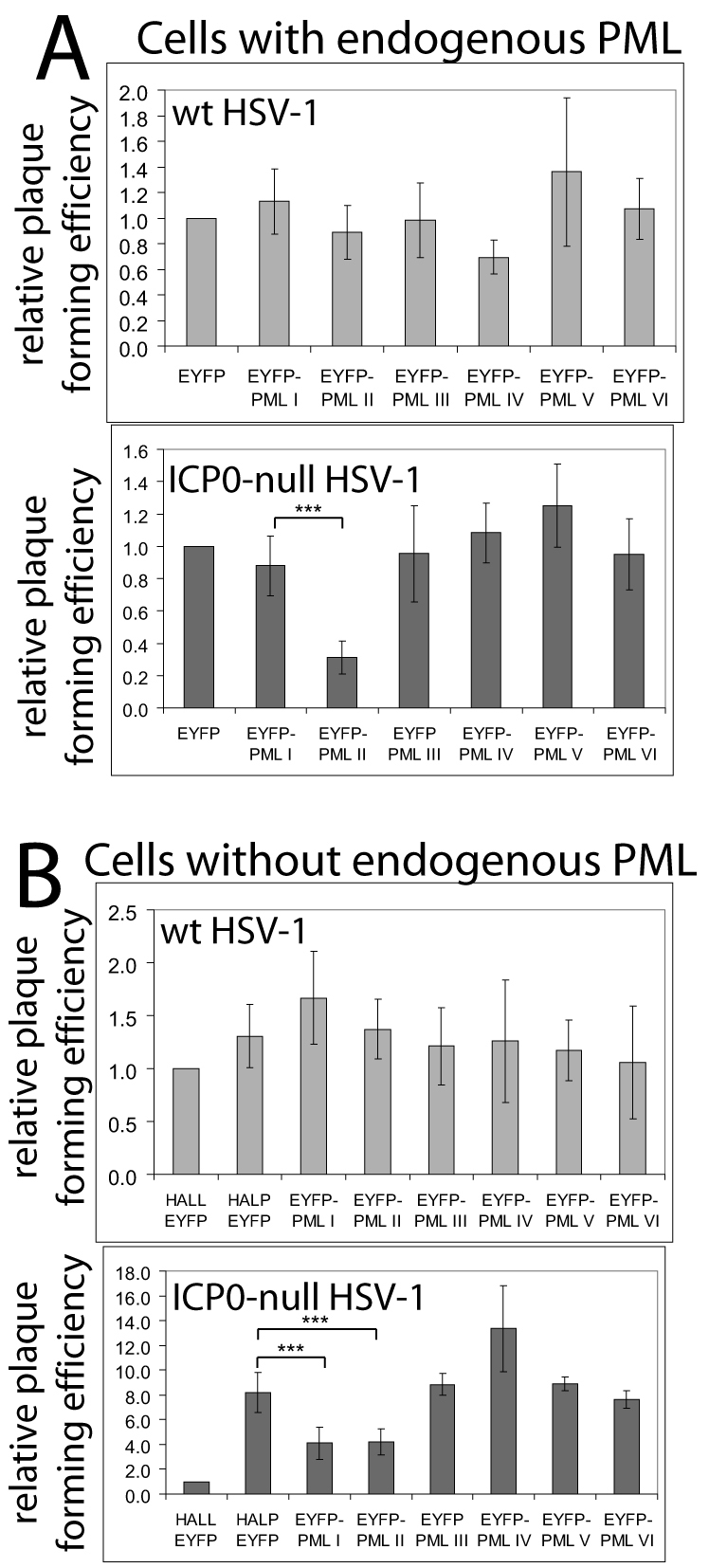
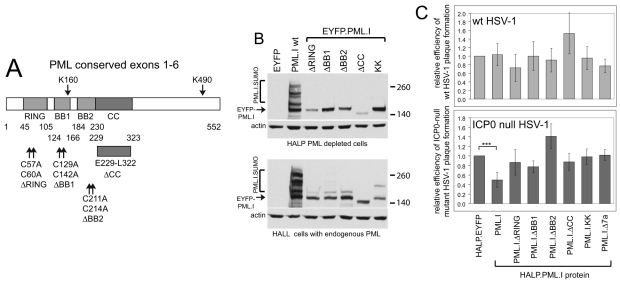
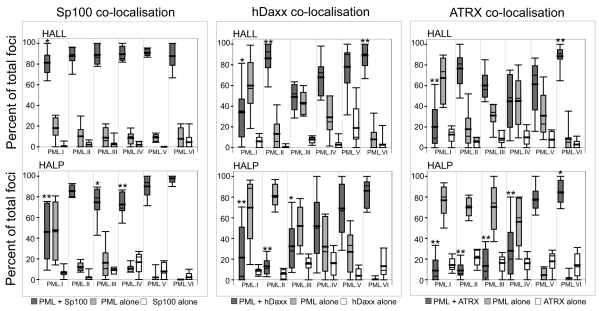
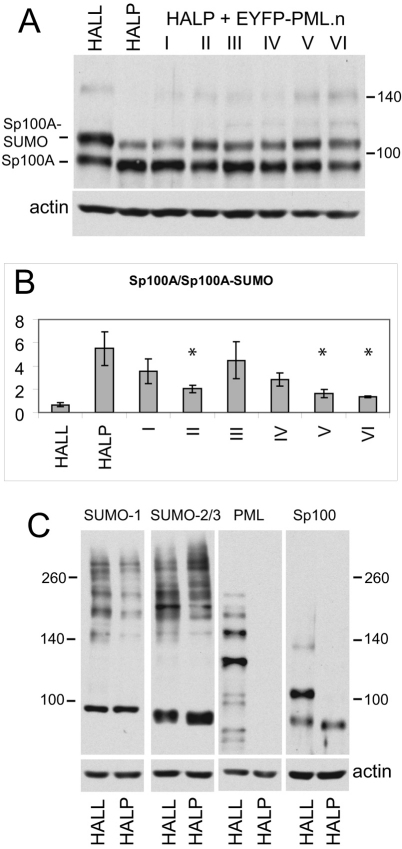
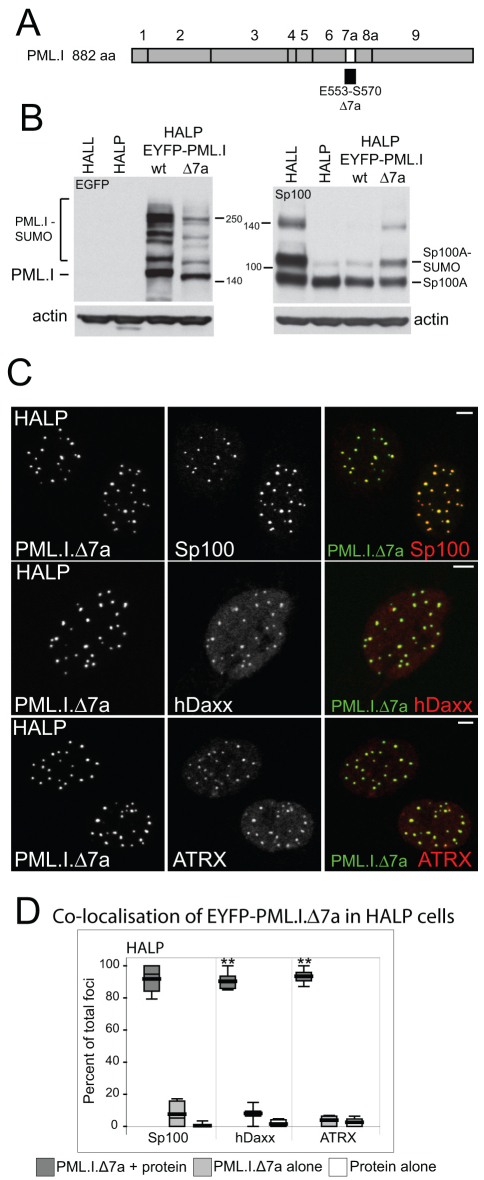
Intrinsic antiviral resistance mediated by constitutively expressed cellular proteins is one arm of defence against virus infection. Promyelocytic leukaemia nuclear bodies (PML-NBs, also known as ND10) contribute to host restriction of herpes simplex virus type 1 (HSV-1) replication via mechanisms that are counteracted by viral regulatory protein ICP0. ND10 assembly is dependent on PML, which comprises several different isoforms, and depletion of all PML isoforms decreases cellular resistance to ICP0-null mutant HSV-1. We report that individual expression of PML isoforms I and II partially reverses the increase in ICP0-null mutant HSV-1 plaque formation that occurs in PML-depleted cells. This activity of PML isoform I is dependent on SUMO modification, its SUMO interaction motif (SIM), and each element of its TRIM domain. Detailed analysis revealed that the punctate foci formed by individual PML isoforms differ subtly from normal ND10 in terms of composition and/or Sp100 modification. Surprisingly, deletion of the SIM motif from PML isoform I resulted in increased colocalisation with other major ND10 components in cells lacking endogenous PML. Our observations suggest that complete functionality of PML is dependent on isoform-specific C-terminal sequences acting in concert.
Figures
References
-
- Beech S. J., Lethbridge K. J., Killick N., McGlincy N., Leppard K. N. (2005). Isoforms of the promyelocytic leukemia protein differ in their effects on ND10 organization. Exp. Cell Res. 307, 109-117 - PubMed
-
- Bernardi R., Pandolfi P. P. (2003). Role of PML and the PML-nuclear body in the control of programmed cell death. Oncogene 22, 9048-9057 - PubMed
-
- Bernardi R., Pandolfi P. P. (2007). Structure, dynamics and functions of promyelocytic leukaemia nuclear bodies. Nat. Rev. Mol. Cell Biol. 8, 1006-1016 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
