

Stress-specific composition, assembly and kinetics of stress granules in Saccharomyces cerevisiae
- PMID: 21172806
- PMCID: PMC3010191
- DOI: 10.1242/jcs.078444
Stress-specific composition, assembly and kinetics of stress granules in Saccharomyces cerevisiae
Abstract
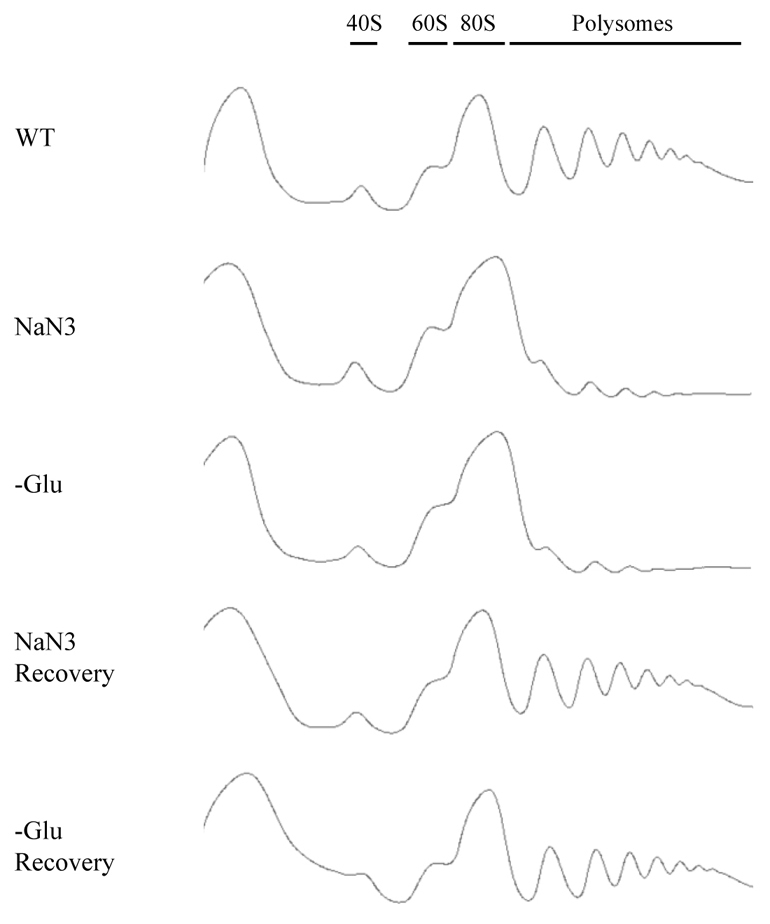
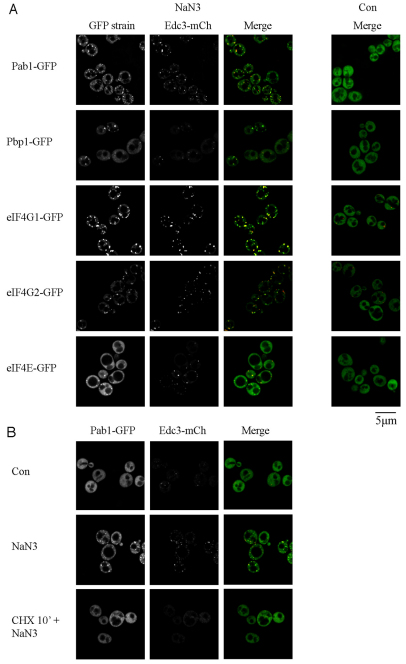
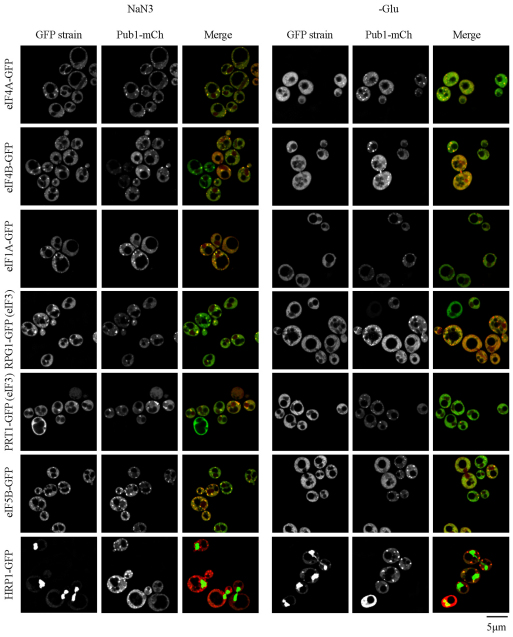
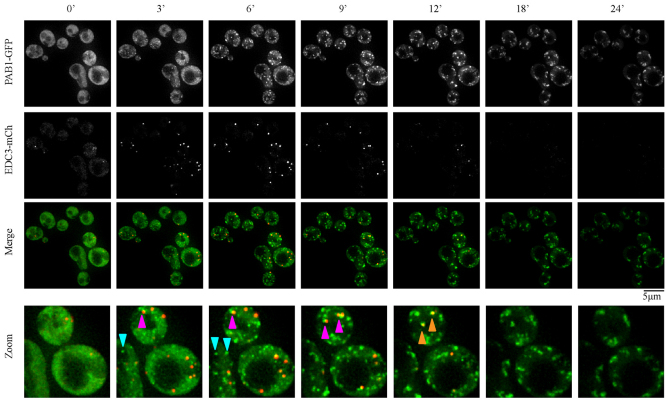
Eukaryotic cells respond to cellular stresses by the inhibition of translation and the accumulation of mRNAs in cytoplasmic RNA-protein (ribonucleoprotein) granules termed stress granules and P-bodies. An unresolved issue is how different stresses affect formation of messenger RNP (mRNP) granules. In the present study, we examine how sodium azide (NaN(3)), which inhibits mitochondrial respiration, affects formation of mRNP granules as compared with glucose deprivation in budding yeast. We observed that NaN(3) treatment inhibits translation and triggers formation of P-bodies and stress granules. The composition of stress granules induced by NaN(3) differs from that of glucose-deprived cells by containing eukaryotic initiation factor (eIF)3, eIF4A/B, eIF5B and eIF1A proteins, and by lacking the heterogeneous nuclear RNP (hnRNP) protein Hrp1. Moreover, in contrast with glucose-deprived stress granules, NaN(3)-triggered stress granules show different assembly rules, form faster and independently from P-bodies and dock or merge with P-bodies over time. Strikingly, addition of NaN(3) and glucose deprivation in combination, regardless of the order, always results in stress granules of a glucose deprivation nature, suggesting that both granules share an mRNP remodeling pathway. These results indicate that stress granule assembly, kinetics and composition in yeast can vary in a stress-specific manner, which we suggest reflects different rate-limiting steps in a common mRNP remodeling pathway.
Figures
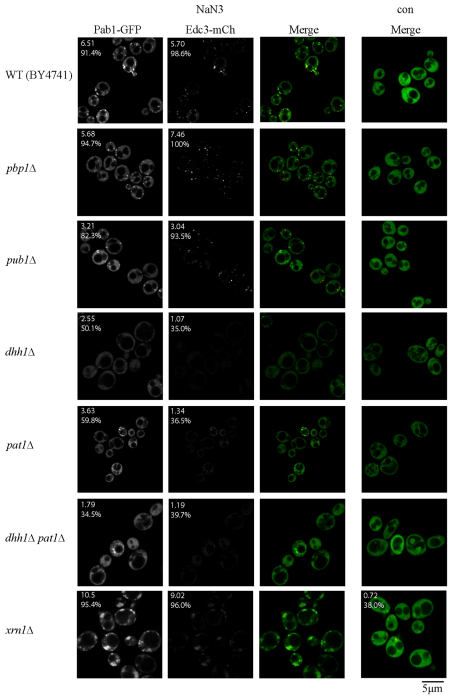
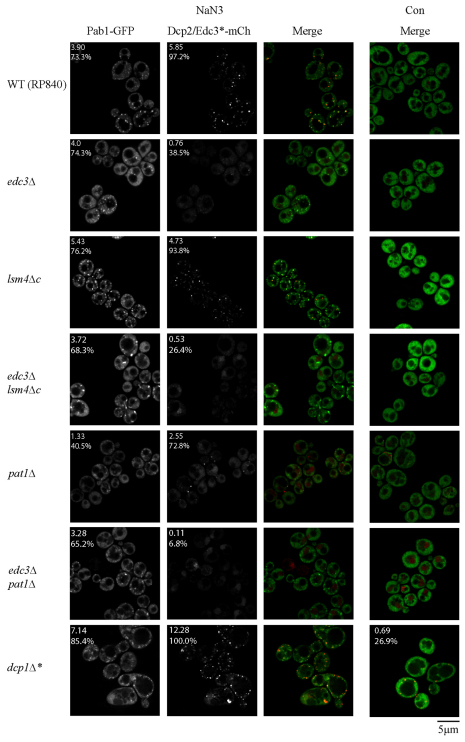
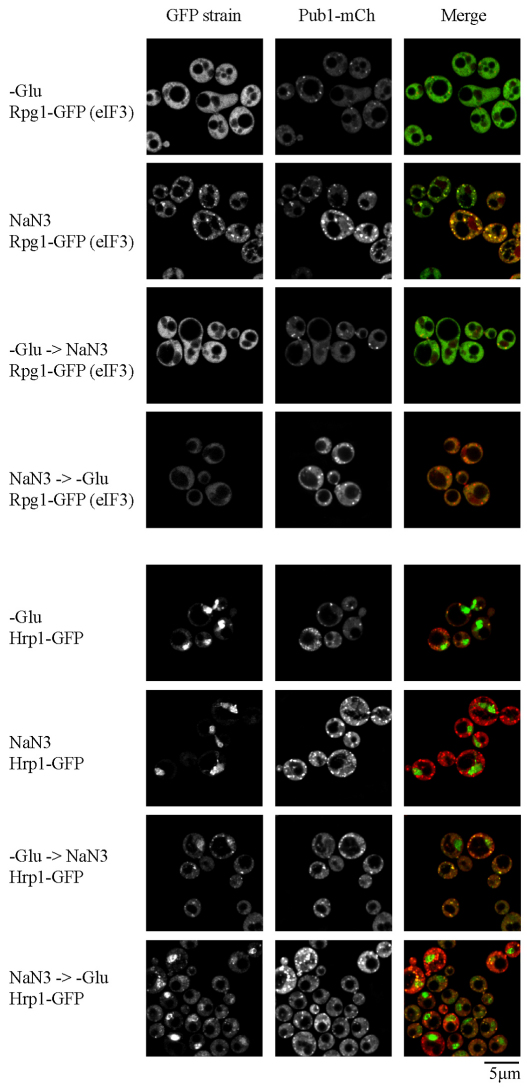
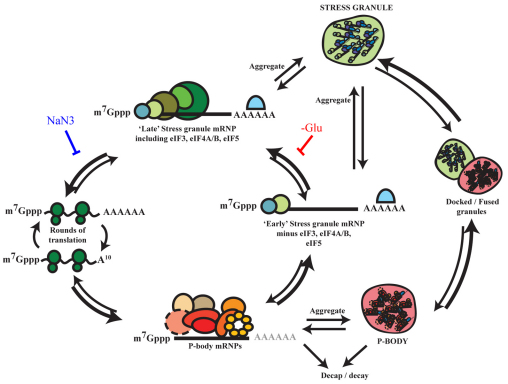
References
-
- Anderson P., Kedersha N. (2009). RNA granules: post-transcriptional and epigenetic modulators of gene expression. Nat. Rev. Mol. Cell Biol. 10, 430-436 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
