

IL-27 promotes T cell-dependent colitis through multiple mechanisms
- PMID: 21173106
- PMCID: PMC3023127
- DOI: 10.1084/jem.20100410
IL-27 promotes T cell-dependent colitis through multiple mechanisms
Abstract
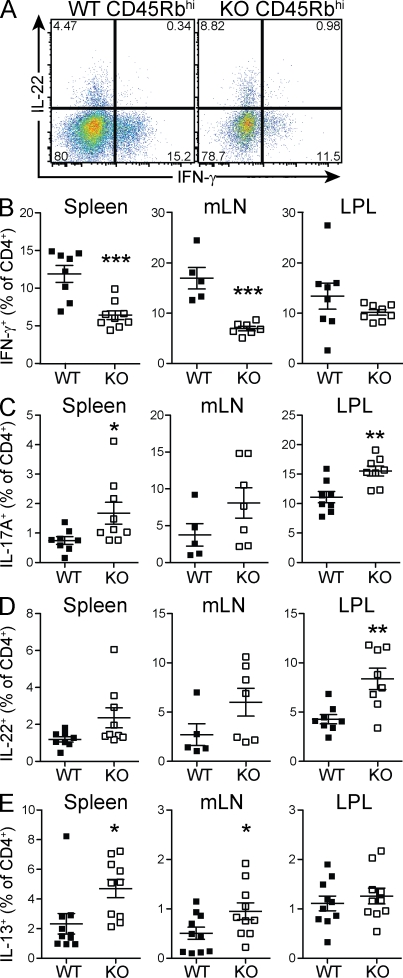
Interleukin-27 (IL-27) is a cytokine known to have both proinflammatory and immunoregulatory functions. The latter appear to dominate in vivo, where IL-27 suppresses TH17 responses and promotes the differentiation of Tr1 cells expressing interferon-γ and IL-10 and lacking forkhead box P3 (Foxp3). Accordingly, IL-27 receptor α (Il27ra)-deficient mice suffer from exacerbated immune pathology when infected with various parasites or challenged with autoantigens. Because the role of IL-27 in human and experimental mouse colitis is controversial, we studied the consequences of Il27ra deletion in the mouse T cell transfer model of colitis and unexpectedly discovered a proinflammatory role of IL-27. Absence of Il27ra on transferred T cells resulted in diminished weight loss and reduced colonic inflammation. A greater fraction of transferred T cells assumed a Foxp3(+) phenotype in the absence of Il27ra, suggesting that IL-27 functions to restrain regulatory T cell (T(reg)) development. Indeed, IL-27 suppressed Foxp3 induction in vitro and in an ovalbumin-dependent tolerization model in vivo. Furthermore, effector cell proliferation and IFN-γ production were reduced in the absence of Il27ra. Collectively, we describe a proinflammatory role of IL-27 in T cell-dependent intestinal inflammation and provide a rationale for targeting this cytokine in pathological situations that result from a breakdown in peripheral immune tolerance.
Figures





Similar articles
-
Interleukin-23 drives intestinal inflammation through direct activity on T cells.Immunity. 2010 Aug 27;33(2):279-88. doi: 10.1016/j.immuni.2010.08.010. Immunity. 2010. PMID: 20732640 Free PMC article.
-
Reciprocal regulation of the survival and apoptosis of Th17 and Th1 cells in the colon.Inflamm Bowel Dis. 2012 Feb;18(2):333-43. doi: 10.1002/ibd.21772. Epub 2011 May 25. Inflamm Bowel Dis. 2012. PMID: 21618360
-
Helicobacter hepaticus-induced colitis in interleukin-10-deficient mice: cytokine requirements for the induction and maintenance of intestinal inflammation.Infect Immun. 2001 Jul;69(7):4232-41. doi: 10.1128/IAI.69.7.4232-4241.2001. Infect Immun. 2001. PMID: 11401959 Free PMC article.
-
CD4+ Foxp3+ regulatory T cells suppress γδ T-cell effector functions in a model of T-cell-induced mucosal inflammation.Eur J Immunol. 2011 Dec;41(12):3455-66. doi: 10.1002/eji.201141814. Epub 2011 Nov 10. Eur J Immunol. 2011. PMID: 21956668
-
Reciprocal IFN-gamma and TGF-beta responses regulate the occurrence of mucosal inflammation.Immunol Today. 1997 Feb;18(2):61-4. doi: 10.1016/s0167-5699(97)01000-1. Immunol Today. 1997. PMID: 9057354 Review.
Cited by
-
Potential clinical application of interleukin-27 as an antitumor agent.Cancer Sci. 2015 Sep;106(9):1103-10. doi: 10.1111/cas.12731. Epub 2015 Aug 6. Cancer Sci. 2015. PMID: 26132605 Free PMC article. Review.
-
Adult-derived human liver stem/progenitor cells as sensors of inflammation: a potential therapy for liver disorders.Hepatobiliary Surg Nutr. 2019 Jun;8(3):286-288. doi: 10.21037/hbsn.2019.01.11. Hepatobiliary Surg Nutr. 2019. PMID: 31245416 Free PMC article. No abstract available.
-
Interruption of macrophage-derived IL-27(p28) production by IL-10 during sepsis requires STAT3 but not SOCS3.J Immunol. 2014 Dec 1;193(11):5668-77. doi: 10.4049/jimmunol.1302280. Epub 2014 Oct 27. J Immunol. 2014. PMID: 25348624 Free PMC article.
-
IL-27R signaling controls myeloid cells accumulation and antigen-presentation in atherosclerosis.Sci Rep. 2017 May 23;7(1):2255. doi: 10.1038/s41598-017-01828-8. Sci Rep. 2017. PMID: 28536468 Free PMC article.
-
Memory CD4 T cells induce selective expression of IL-27 in CD8+ dendritic cells and regulate homeostatic naive T cell proliferation.J Immunol. 2012 Jan 1;188(1):230-7. doi: 10.4049/jimmunol.1101908. Epub 2011 Nov 23. J Immunol. 2012. PMID: 22116827 Free PMC article.
References
-
- Artis D., Villarino A., Silverman M., He W., Thornton E.M., Mu S., Summer S., Covey T.M., Huang E., Yoshida H., et al. 2004. The IL-27 receptor (WSX-1) is an inhibitor of innate and adaptive elements of type 2 immunity. J. Immunol. 173:5626–5634 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
